



Ультраструктура парных маутнеровских нейронов при вестибулярной или оптокинетической стимуляции и предварительной аппликации β-амилоида
Моделирование деструктивных процессов в нейронах при изучении морфофункциональных аспектов нейродегенеративных заболеваний предполагает использование конкретных нейронов с установленной функцией. Такими, на наш взгляд, являются маутнеровские нейроны (МН), две идентифицированные клетки головного мозга костистых рыб. Благодаря своей величине, строению и зеркальному расположению в продолговатом мозге, их безошибочно определяют на гистологических срезах [6, 7]. Афферентные входы к МН со стороны статоакустического и зрительного аппарата образуют контакты с соматической частью и основными дендритами – латеральным и вентральным. Эти нейроны рассматривают в качестве главного элемента сенсомоторной сети, управляющей латеральной формой локомоции – совершением поворотов при плавании, когда левый нейрон инициирует поворот вправо, а правый – влево [10]. При этом у рыбок «правшей» доминантным является левый нейрон, а у рыбок «левшей» – правый. Нами была разработана экспериментальная клеточная модель амилоидоза с аппликацией на МН бета-амилоида, основного белка сенильных бляшек, морфологического индикатора болезни Альцгеймера [7]. Под действием этого белка происходила инверсия латерального поведения золотых рыбок, в основе которого – дистрофия МН, нарушение их трехмерного строения и структурной асимметрии между нейронами [1]. Длительные сенсорные стимуляции в сочетании с аппликациями бета амилоида приводили к тому, что деформированные нейроны могли образовывать дополнительный медиальный дендрит, никогда ранее не наблюдавшийся у МН [2,3]. В связи с этими данными, представляется необходимым исследовать эффекты вестибулярной и зрительной стимуляции, активирующей разные входы к МН на нейродегенеративные процессы в каждом нейроне из пары и их афферентных синапсах, индуцированные применением бета-амилоида.
Материал и методы исследования
В работе использовали МН мальков золотой рыбки (CarassiusauratusL.) породы Оранда 4 мес возраста. Условия содержания рыбок, определение их моторной латерализации, аппликации агрегированного фрагмента 25–35 бета-амилоида (далее βА25-35) в дозе 20 мкг в 4 мкл дистиллированной воды, проведение спустя 24 час длительной вестибулярной стимуляции (ротации рыбок одновременно вокруг длинной и короткой осей тела в течение 2 час) в деталях опубликовано ранее [2, 3]. Оптокинетическую стимуляцию в течение 10 час, основанную на инстинктивном следовании рыбок за перемещающимися зрительными ориентирами в направлении противоположном естественной латерализации рыбок, проводили спустя 17 час. Такое воздействие направлено на стимуляцию доминантного нейрона из пары [5].
Для морфологического исследования рыбок декапитировали, выделяли участок продолговатого мозга, содержащий оба нейрона, фиксировали поэтапно в глутаральдегидном фиксаторе на какодилатном буфере с добавлением формальдегида и диметилсульфоксида и в 1 % растворе четырехокиси осмия, обезвоживали в спиртах возрастающей концентрации и абсолютном ацетоне, заключали в эпон. С эпоновых блоков ткани мозга каждой рыбки с помощью пирамитома (LKB, Швеция) изготовляли серийные гистологические срезы толщиной 3 мкм с четкой идентификацией левого и правого нейрона в каждом срезе. С помощью методики 3D реконструкции МН получали их объемные изображения, по которым анализировали морфологические изменения. Ультраструктурному исследованию подвергали только те парные нейроны, один из которых формировал дополнительный медиальный дендрит. Для проведения электронно-микроскопического исследования гистологические срезы переклеивали на новые эпоновые блоки, резали на ультратоме EMUC6 (Leica, ФРГ). Контрастировали стандартно уранилацетатом и цитратом свинца, изучали в электронном микроскопе TeslaBS-500 при увеличении х14 тыс. Подробно методическая часть работы по приготовлению препаратов для прицельного совмещенного светового и электронно-микроскопического исследования и физиологическая часть работы с морфологическим исследованием были опубликованы ранее [2, 3, 4,].
Результаты исследования и их обсуждение
1. Исследование ультраструктуры МН после аппликации βА25-35 и вестибулярной стимуляции рыбок показало, что, как и ожидалось, нейроны проявляют признаки дистрофических изменений. В частности, ультраструктурный анализ правого и левого нейронов у одной из опытных рыбок выявил изменения, характерные для последствий вестибулярной стимуляции, но в условиях действия βА25-35 они становятся еще более выраженными. Нейроны уменьшаются в размерах, что отмечалось ранее, но сохраняют асимметрию объемов: один нейрон больше другого [2]. Правый нейрон, по функциональным данным, определяемым на основании моторной латерализации, в данном случае рыбки-левши, до начала экспериментов был доминантным, т.е. более активным. По окончании эксперимента в функциональном отношении оба нейрона становятся равнозначными. При этом у меньшего по размеру правого нейрона сильно выражены признаки истощения и деструкции. Плазматическая мембрана местами не определяется. Ее извилистый контур образует складки и узкие инвагинации в пространство между боковыми профилями аксональных отростков, образующих синаптические контакты с МН (рисунок, А). Такие инвагинации подобны тем, которые наблюдаются при хроническом воздействии этанолом, меняющим физические свойства клеточных мембран [9]. Профиль ядра становится вытянутым в медио-латеральном направлении. Диффузный хроматин (у интактных МН) конденсируется в глыбки, в кариоплазме появляются кристаллические структуры. Такой эффект, как правило, наблюдается у МН, при утомлении их функции после длительной вестибулярной стимуляции [6]. Цитоплазма в соматической части и в вентральном дендрите электронноплотная. Цитоскелет плохо дифференцируется: еле различимые нити нейрофиламентов перемежаются областями, заполненными свободными рибосомами (рисунок, А). В соматической части и в проксимальных отделах дендритов в постсинаптических областях встречаются ячеистые включения. Считается, что образование таких структур связано с грубыми нарушениями структуры цитоплазматических белков [6]. Меняется строение митохондрий. При полной сохранности формы этой органеллы и ее наружной мембраны заметна редукция крист с образованием в центре митохондрии либо вакуоли, либо электронно-плотного округлого образования, либо миелиновой структуры. Деструкция митохондрий обнаруживается как в соматической части, так и в вентральном дендрите. Отдельные большие миелиновые структуры, характерный признак последствий вестибулярной стимуляции, встречаются преимущественно в субсинаптической области.
Левая клетка, большая по размеру, образующая медиальный дендрит, претерпевает существенно меньшие деструктивные изменения (рисунок, Б). Ядро содержит гомогенный хроматин, сохраняет округлую форму, образует узкие длинные инвагинации в цитоплазму. Ядерная оболочка в инвагинирующих участках насыщена порами. Цитоплазма в соматической части и в медиальном дендрите менее электронноплотная, но вентральный дендрит все же остается более осмиофильным. Цитоскелет определяется достаточно хорошо. Нейрофиламенты находятся на большем расстоянии друг от друга и сохраняют параллельность расположения. Рибосомы преимущественно собраны в полисомы, эндоплазматический ретикулум слабо гипертрофирован. Митохондрии несколько набухшие, кристы расположены по периферии.
Афферентный синаптический аппарат обоих МН в морфологическом отношении представлен синапсами химического и смешанного типа (рисунок, А, Б).
Признаки дегенеративных изменений отмечаются у небольшого количества синапсов химического типа, образующих контакты в виде активных зон или активной зоны в соседстве с десмосомоподобной структурой как с соматической частью МН, так и с дендритами. Такие бутоны имеют больший размер и незначительное количество синаптических везикул у активной зоны. Характерным патологическим признаком является наличие у них вакуолей и мембранных структур. Редко встречаются синаптические бутоны с аглютинированными везикулами или опустошенные бутоны. При этом митохондрии в синапсах сохраняют кристы во всем объеме органеллы. Особо следует подчеркнуть сохранность синаптических контактов. Все три типа специализированных контактов: активные зоны, щелевые и десмосомоподобные контакты идентифицируются достаточно хорошо.
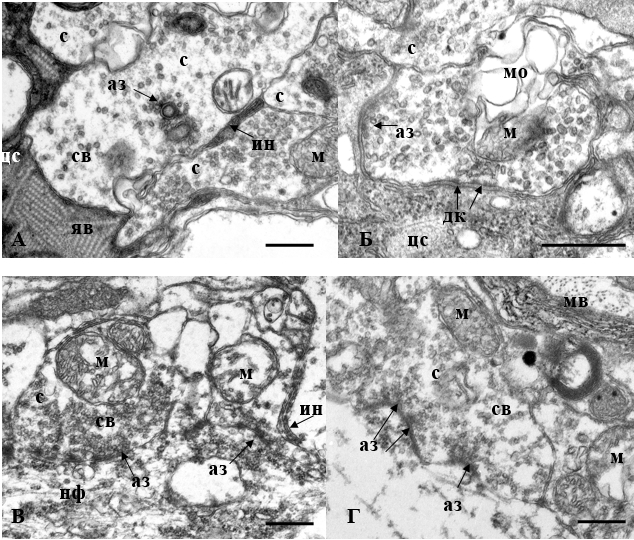
Ультраструктура маутнеровских нейронов после аппликации на них фрагмента 25–35 бета-амилоида и вестибулярной (А, Б) или оптокинетической (В, Г) стимуляции золотой рыбки
А – изменения в правом нейроне: появление инвагнаций в области соседствующих аксонных окончаний (ин), уплотнение цитоскелета (цс), образование ячеистых включений в цитоплазме (яв). Некоторые митохондрии (м) в синапсах (с) сохраняют кристы. Представлены гребешковые (шипикоподобные) синапсы с активной зоной (аз) на гребешке.
Б – изменения в левом нейроне: электронноплотная цитоплазма, элементы цитоскелета слабо дифференцированы, в бутоне видны мембранные образования (мо). Специализированные контакты – активные зоны и десмосомоподобные структуры (дк) – определяются хорошо.
В – изменения в правом нейроне: цитоплазма электронносветлая, из элементов цитоскелета присутствуют короткие фрагменты нейрофиламентов (нф), инвагинация плазматической мембраны нейрона в область между аксонными окончаниями.
Г – изменения в левом нейроне: деструкция нейрона, агглютинация синаптических везикул не только у предполагаемой активной зоны, но и во всем объеме бутона, разрывы плазматической мембраны нейрона, локальное расслоение и извитость контуров мембранных витков (мв) миелиновой оболочки аксона.
Масштабный отрезок – 0.5 мкм.
2. После аппликации βА25-35 и зрительной стимуляции признаки деструкции МН выражены в большей степени, чем при вестибулярной стимуляции. У рыбки-левши при инверсии моторной латерализации вследствие оптокинетической стимуляции изначально доминантный правый нейрон становится субдоминантным, менее активным. Он большего размера, чем левый нейрон информирует медиальный дендрит при сильной дистрофии вентрального дендрита. Плазматическая мембрана этого нейрона теряет четкость контура. Цитоплазма становится электроннопрозрачной. В существенной степени дезорганизован цитоскелет, как было отмечено ранее [3]. Параллельное расположение редких нейрофиламентов нарушается. Другой элемент цитоскелета, актиновые нити, отсутствуют (рисунок, В). Матрикс митохондрий просветлен, кристы оттеснены к периферии органеллы, центр может содержать вакуоль. Миелиновые структуры не обнаруживаются. В вентральном дендрите цитоплазма менее прозрачна, параллельное расположение нейрофиламентов не нарушается, их заметно больше, но между ними также большие промежутки. Синаптические окончания сохраняют кластерное расположение, при этом встречаются кластеры опустошенных бутонов и кластеры бутонов, заполненных мелкими везикулами (рисунок, В). Такими чаще всего бывают гребешковые синапсы. В заполненных бутонах везикулы аглютинируются в глыбки. Строение синаптических везикул нарушается: их мембрана истончается, теряет четкость, видны отходящие тончайшие фибриллы. Из специализированных синаптических контактов относительно хорошо определяются десмосомоподобные и щелевые контакты, активные зоны идентифицируются хуже.
Другой нейрон из этой пары, левый, ставший доминантным после интенсивной функциональной нагрузки, подвергся сильной деструкции (рисунок, Г). Он значительно меньше по объему, сформированный им медиальный дендрит также мал, как и другие его дендриты. При ультраструктурном рассмотрении плазматическая мембрана прослеживается не везде и имеет разрывы. Просветление цитоплазмы сопровождается появлением значительных областей, не содержащих органелл (рисунок, Г). Из элементов цитоскелета представлены только редко встречающиеся нейрофиламенты. Сильно поврежден афферентный синаптический аппарат. Значительное количество бутонов опустошается, а оставшиеся синаптические везикулы сосредоточены только в одном локальном месте, предполагаемой активной зоне. Встречаются бутоны, заполненные патологически измененными везикулами. У расположенных рядом миелиновых волокон заметна очаговая деформация мембранных витков (рисунок, Г).
Заключение
Сравнительное изучение ультратонкого строения парных МН опытных золотых рыбок позволяет сделать следующие выводы.
1. Длительные сенсорные нагрузки усиливают деструктивное действие бета-амилоида на нейроны. Изменения преимущественно касаются нарушения нормального строения нейронального цитоскелета, митохондрий и афферентного синаптического аппарата нейронов. Характер нейродегенеративных процессов различен и зависит от типа воздействия, активирующего определенный афферентный вход к МН. При оптокинетической стимуляции деструктивные явления выражены в большей степени, чем при вестибулярной стимуляции вплоть до полной деструкции органелл. Причина различий, на наш взгляд, связана не только со специфичностью воздействия, но и с его продолжительностью.
2. Парные МН одной особи проявляют разную степень деструкции при стимуляции одним и тем же раздражителем. У одного нейрона из пары признаки дегенерации выражены сильнее, чем у другого. Вероятно, это является результатом более сильной физиологической нагрузки на нейрон, который является (или становится) функционально доминантным (более активным) в данной паре. Не однозначна также роль сформированного клетками медиального дендрита. Выяснение причины образования такого дендрита и его значимости требует отдельного рассмотрения.
3. Результаты исследования позволяют заключить, что МН и аппликации на них бета-амилоида могут служить адекватной моделью для изучения нейродегенерации и возможных путей защиты индивидуальных нейронов от повреждений, вызванных токсическими воздействиями и утомляющими стимуляциями.
Библиографическая ссылка
Тирас Н.Р., Михеева И.Б., Пенькова Н.А. ИЗМЕНЕНИЯ В УЛЬТРАСТРУКТУРЕ ПАРНЫХ МАУТНЕРОВСКИХ НЕЙРОНОВ ЗОЛОТОЙ РЫБКИ, СВЯЗАННЫЕ С ВЕСТИБУЛЯРНОЙ И ОПТОКИНЕТИЧЕСКОЙ СТИМУЛЯЦИЕЙ И АППЛИКАЦИЕЙ НА НЕЙРОНЫ БЕТА-АМИЛОИДА // Современные проблемы науки и образования. – 2016. – № 6. ;URL: https://science-education.ru/ru/article/view?id=25562 (дата обращения: 18.04.2024).