



Лактатдегидрогеназа (ЛДГ) – это один из ключевых ферментов гликолиза, который, находясь на развилке путей метаболизма углеводов, участвует в регуляции тонко сбалансированного анаэробного и аэробного гликолиза. Однако функционирование ЛДГ может быть связано не только с его центральной ролью в рециркуляции анаэробного гликолиза, но и в регуляции соотношения НАД+/ НАДН, поскольку именно оно влияет на скорость многих каталитических реакций. Было показано, что внутриклеточный НАД+ играет важную роль в регуляции транскрипции генов, связанных с метаболизмом и циркадианными ритмами [1]. Предполагается, что ЛДГ может участвовать в клеточном цикле [2], в регуляции активности АТФ зависимых K+ каналов [3].
Все многообразие внутриклеточных процессов, связанных определенным образом с каталитической активностью ЛДГ, предполагает необходимость изучения молекулярных механизмов функционирования этого фермента при различных физиологических состояниях животных, в частности при гипотермии. В последние годы гипотермические состояния гомойотермных животных привлекают особый интерес, поскольку их все чаще применяют для повышения устойчивости органов и тканей к различным видам стресса, защиты организма от травматических повреждений, восстановления функции органов и тканей после ишемии-реперфузии, для коррекции и лечения различных заболеваний животных и человека [4].
Ранее нами было показано, что при умеренной гипотермии у крыс происходят существенные изменения активности и кинетических параметров ЛДГ мозга и скелетных мышц [5; 6]. При этом каталитические свойства ЛДГ печени, являющейся интегратором всех биохимических процессов организма в целом, до сих пор остаются неизвестными.
Материалы и методы исследования
Исследования проводили на белых беспородных крысах (самцах) массой 150-200 г. Умеренную гипотермию вызывали в холодовой камере, в рубашке которой циркулировала вода температурой 4-7 ºС. Температуру тела животных в течении 30 мин доводили до 30 ºС.
Получение тканевого экстракта. Животных декапитировали, выделяли печень и гомогенизировали ее. Гомогенат центрифугировали при 600 g в течение 10 мин. Полученный супернатант повторно центрифугировали при 15000 g в течение 10 мин. Содержание белка в полученном экстракте (безмитохондриальном цитозоле) определяли по методу Лоури [7].
Определение активности ЛДГ. Активность ЛДГ определяли по убыли содержания НАДН в реакционной смеси, что регистрировалось спектрофотометрически. Реакционная смесь содержала 2.4 мл 0.1 М фосфатного буфера (рН 7.4), 0.3 мл раствора пирувата натрия (Sigma, США), 0.3 мл 1 мМ раствора НАДН (Sigma, США) и 0,05 мл тканевого экстракта. Исследование активности ЛДГ проводили в диапазоне концентраций пирувата от 0.05 до 25.6 мМ. Активность ЛДГ выражали в наномолях НАДН, окисленного в результате ферментативной реакции за 1 мин на 1 мг белка (нмоль/мин·мг белка).
Определение кинетических характеристик. Кинетические характеристики (максимальную скорость (Vm), константу Михаэлиса (Кm) и константу ингибирования (Ki)) вычисляли с помощью программного пакета Statistica 6.0 посредством нелинейного многомерного регрессионного анализа, используя в опции «нелинейное оценивание» уравнение Холдейна.
Статистическая обработка результатов. Обработка данных произведена с помощью однофакторного дисперсионного анализа (ANOVA) с использованием пакета Statistica 6.0. Достоверность различия определяли с помощью критерия Фишера на уровне значимости P=0.05. Каждая кривая на графиках концентрационной зависимости скорости окисления НАДН - среднее восьми независимых экспериментов. Данные в таблице приведены в виде: среднее ± ошибка среднего.
Результаты исследования и их обсуждение
Была исследована концентрационная зависимость ЛДГ печени крыс в норме. Результаты исследования представлены на рис. 1. Из рис. 1 видно, что кривая концентрационной зависимости ЛДГ печени крыс имеет колоколообразный характер, указывающий на то, что активность фермента увеличивается с повышением концентрации субстрата до максимального значения при определенной концентрации субстрата, называемой точкой оптимума. Эта концентрация для печени интактных крыс составляет ≈0,5 мМ. Дальнейшее повышение концентрации субстрата приводит к снижению активности ЛДГ, что свидетельствует о наличии у фермента феномена субстратного ингибирования. Ранее сотрудниками кафедры биохимии и биофизики субстратное ингибирование ЛДГ было обнаружено в тканях мышц и мозга крыс [5; 6]. Субстратное ингибирование обусловлено формированием и диссоциацией ковалентного аддукта между пируватом и окисленной формой кофактора [8]. Остаток аминокислоты серина-163 играет ключевую роль в этом механизме, так же как присутствие нескольких активных центров и взаимодействие между субъединицами в тетрамерном белке.
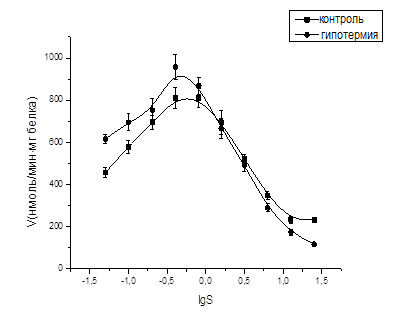
Рис. 1. Концентрационная зависимость активности ЛДГ печени крыс в норме и при гипотермии
Интересно то, что правый (ингибиторный) склон концентрационной зависимости ЛДГ демонстрирует сигмоидный характер зависимости от концентрации пирувата. Это является свидетельством того, что при некоторой критической концентрации пирувата происходит резкая смена характера ингибирования ЛДГ.
Из рис. 1 видно, что гипотермия приводит к повышению активности ЛДГ в диапазоне низких концентраций пирувата (0,05-0,8 мМ). Так, например, повышение активности ЛДГ при концентрации 0,4 мМ составляет 19,2%. Интересно то, что в области высоких концентраций пирувата (0,8-25,6мМ) гипотермия, напротив, сопровождается снижением активности фермента. Такое снижение становится наиболее выраженным при максимально использованной концентрации пирувата (25,6 мМ), составляя 50,1%.
Рис. 1. наглядно демонстрирует, что гипотермия влияет не только на активность фермента, но и на характер его концентрационной зависимости: точка оптимума на концентрационной кривой смещается в сторону более низких концентраций пирувата. Кроме того, сигмоидность на правом ингибиторном склоне графика концентрационной зависимости становится менее выраженной. Вместе с тем сигмоидность появляется на левом активаторном склоне концентрационной кривой, что свидетельствует о триггерном механизме связывания субстрата в определенном узком диапазоне.
Поскольку лактатдегидрогеназная реакция характеризуется наличием субстратного ингибирования, то для описания зависимости скорости реакции от концентрации пирувата неприемлемо уравнение Михаэлиса-Ментэн. Субстратное ингибирование для односубстратных ферментативных реакций хорошо описывается моделью Холдейна. Однако в лактатдегидрогеназной реакции участвует не только пируват, но и кофактор - НАДН. Таким образом, эту реакцию можно отнести к двусубстратным, и тогда применение модели Холдейна также представляется неадекватным. Однако, учитывая тот факт, что исследование проводили при фиксированных насыщенных концентрациях кофактора, эффектами его на скорость ферментативной реакции можно пренебречь и использовать уравнение Холдейна в качестве регрессионной модели.
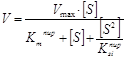 ;
;
где Vmax – максимальная скорость обратимой реакции;
Km – константа Михаэлиса для пирувата;
Ksi – константа субстратного ингибирования для пирувата;
[S] – концентрация пирувата.
Используя это уравнение в опции «нелинейное оценивание» с помощью нелинейного регрессионного анализа были вычислены кинетические характеристики лактатдегидрогеназы (рис. 2, таблица). Исследование показало, что при гипотермии показатель Vm незначительно (на 14, 9%) увеличивается. При этом Кm уменьшается на 20,1%. Незначительное повышение Vm на фоне более существенного повышения Кm приводит к тому, что при гипотермии на 43,2% повышается эффективность катализа – величина, характеризуемая отношением Vm/Km. Эффективность катализа численно равна константе скорости первого порядка и отражает скорость ферментативной реакции при физиологических концентрациях субстрата.
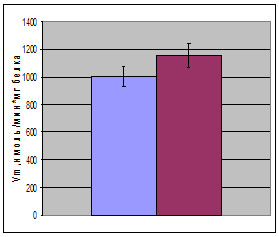
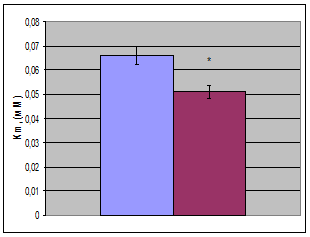
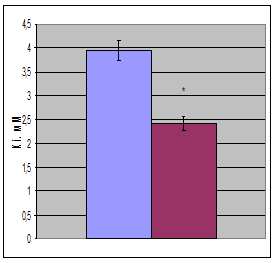
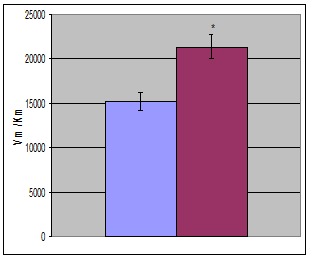
Рис. 2. Кинетические характеристики (Кm, Vm, Vm/Km, Ki) ЛДГ печени крыс в норме и при умеренной гипотермии (  - контроль,
- контроль,  - умеренная гипотермия); *- достоверность различий относительно контроля
- умеренная гипотермия); *- достоверность различий относительно контроля
Исследование показало, что гипотермия оказывает существенное влияние на параметры субстратного ингибирования фермента. Из рис. 2 видно, что значение Кi при гипотермии значительно (на 38,7%) снижается. Это указывает на то, что при гипотермии увеличивается сродство пирувата, при его высоких концентрациях, к активному центру фермента. Это демонстрирует то, что для ингибирования ЛДГ при гипотермии требуются более низкие концентрации пирувата, нежели в контроле. Такое снижение константы ингибирования ЛДГ при гипотермии приводит к тому, что в 2,4 раза увеличивается значение коэффициента субстратного ингибирования (Q), определяемого нами как отношение скорости катализа при оптимальной концентрации субстрата к скорости катализа при максимально взятой в данной работе концентрации субстрата.
Кинетические характеристики ЛДГ мозга крыс в норме и при гипотермии
|
Состояние животного |
Q |
Sопт |
?=Кi - Кm |
|
контроль |
3,51±0,31 |
0,51±0,04 |
3,877±0,02
|
|
гипотермия |
8,26±0,62* |
0,36±0,02* |
2,365±0,03*
|
* - достоверность различий относительно контроля.
Поскольку гипотермия приводит к снижению значений Кm и Ki , это в совокупности способствует снижению значения оптимальной концентрации субстрата (Sopt), что выражается в сдвиге точки максимума на графике концентрационной зависимости в сторону более низких концентраций пирувата. В соответствии с моделью Холдейна можно вычислить точку оптимума. Вычисленные нами теоретические значения Sopt примерно соответствуют графическим и демонстрируют снижение значения Sopt при гипотермии на 29,5%. Нами была вычислена разница между значениями Ki и Km (?=Ki-Km), характеризующая диапазон концентрации пирувата, при котором фермент работает с максимальной эффективностью. Оказалось, что при гипотермии этот диапазон уменьшается на 39%.
Таким образом, исследование показало, что при гипотермии происходят существенные изменения многих кинетических характеристик ЛДГ. При этом повышение максимальной скорости катализа ЛДГ не является достоверным. Максимальная скорость – это величина, которая определяется произведением числа оборотов фермента и его концентрации. Концентрация фермента может изменяться вследствие изменения соотношения скоростей его синтеза и распада. Синтез фермента – это энергоемкий процесс. При гипотермии, в условиях снижения скоростей всех энергосинтезирующих процессов и энергопотребляющих процессов, экспрессия многих белков подавлена. Учитывая, что в печени, как и во всех других органах, могут быть представлены различные изоформы ЛДГ, среди которых наиболее высока концентрация ЛДГ 4 и ЛДГ 5, можно предположить, что при гипотермии происходят изменения не столько количества фермента, сколько его «качества», то есть изменяется количественное соотношение экспрессируемых изоформ ЛДГ. Однако в условиях низких температур возможность дифференциальной экспрессии генов, кодирующих различные изозимы, тоже, скорее всего, будет значительно снижена.
Таким образом, временные рамки гипотермии и энергодефицит исключают возможность количественной регуляции активности ЛДГ. Поскольку в нашем исследовании более значимые изменения при гипотермии обнаружены в значениях константы Михаэлиса, то это указывает на то, что повышение эффективности катализа связано с посттрансляционными модификациями в самой молекуле фермента. О механизмах регуляции ЛДГ гомойотермных животных посредством посттрансляционной модификации известно мало. При этом большое разнообразие механизмов регуляции ЛДГ обнаружено у пойкилотермов: фосфорилирование, аденилирование, ацетилирование, убиквитирование и т.д. [9].
Имеются единичные экспериментальные данные, указывающие на регуляцию ЛДГ в клетках опухолевых линий млекопитающих посредством фосфорилирования по остаткам тирозина [10]. В регуляции ЛДГ могут принимать участие шапероны, которые могут изменять конформацию фермента таким образом, что сродство его к субстрату будет значительно увеличиваться. Обнаружено, что при гипотермии изменяется уровень экспрессии шаперонов в тканях гомойотермов [11]. Одной из причин модификации структуры ЛДГ может стать её окисление АФК, концентрация которых многократно увеличивается при стрессе. Было обнаружено, что гидроксильные радикалы, генерируемые in vitro в среде Фентона, приводят к угнетению активности ЛДГ [12]. Как известно, гипотермические состояния сопровождаются активацией свободно-радикальных процессов [13]. Поэтому гипотеза об окислительной модификации ЛДГ под действием активных форм кислорода кажется вполне правдоподобной.
Повышение эффективности катализа ЛДГ в результате снижения значения Кm указывает на реализацию у гомойотермных животных в условиях низких температур механизмов температурной компенсации. Температурная компенсация – это явление, характерное для пойкилотермов и позволяющее поддерживать высокие скорости ферментативных реакций при низких температурах тела животного [14].
Наши данные указывают на реализацию у гомойотермных животных в условиях низких температур механизмов температурной компенсации. Возможно, что в процессе эволюции гомойотермных животных, несмотря на формирование у них принципиально новой стратегии адаптации к температуре, в зачаточном состоянии сохраняются элементы пойкилотермных механизмов регуляции активности фермента. Эти механизмы реализуется в период экстремальных температур окружающей среды, когда терморегуляционные системы уже не справляются с поддержанием постоянной температуры тела.
Библиографическая ссылка
Халилов Р.А., Джафарова А.М., Хизриева С.И., Абдуллаев В.Р. КИНЕТИЧЕСКИЕ СВОЙСТВА ЛАКТАТДЕГИДРОГЕНАЗЫ ПЕЧЕНИ КРЫС В НОРМЕ И ПРИ УМЕРЕННОЙ ГИПОТЕРМИИ // Современные проблемы науки и образования. – 2017. – № 5. ;URL: https://science-education.ru/ru/article/view?id=27087 (дата обращения: 26.04.2024).