



Основные задачи настоящего исследования были связаны с изучением механизмов действия различных доз γ-радиации на рост растения. Известно, что γ-облучение в малых стимулирующих дозах приводит к увеличению всхожести и энергии прорастания семян, ускорению роста и развития, процессов дыхания и фотосинтеза растений, и, в конечном итоге, к повышению их урожайности. Это происходит за счет сдвигов в метаболизме облученных прорастающих семян уже на самых ранних стадиях развития - ускоренной мобилизации питательных веществ, интенсификации окислительных процессов, изменения нуклеотидного состава РНК и ДНК [2, 3]. Под действием высоких, угнетающих доз γ-облучения происходит замедление роста, появление уродливых форм и последующая гибель растений. Это происходит за счет образования свободных радикалов, уменьшения содержания нуклеиновых кислот, мутаций ДНК, окислительного стресса - значительной активации окислительных процессов, затрагивающих и ферментные системы. Торможение роста также связывалось с ингибирование синтеза и содержания ауксинов - физиологических фитогормонов роста, однако не были окончательно выяснены механизмы действия излучений на синтез ауксинов, в частности, ИУК, способствующей росту клеток путем их растяжения [8,12]. Целью настоящей работы было исследование влияния γ-радиации на процессы биосинтеза ИУК из индольных производных в проростках, полученных из облученных семян, а также на некоторые показатели азотного обмена [5].
Были исследованы также роль и механизм действия различных концентраций ИУК на мембраны растительной клетки. Известно, что одним из механизмов действия ИУК в клетке является активация транспорта ионов водорода (Н+) через клеточные цитоплазматические мембраны [7]. Представляло интерес исследование действия ИУК на других типах биологических мембран (тилакоидов и модельных БЛМ). Согласно хемиосмотической гипотезе П.Митчела [17], преобразование энергии квантов света в энергию химических связей аденозинтрифосфорной кислоты (АТФ) в мембранах тилакоидов происходит в результате транспорта электронов по ЭТЦ, сопряженного с переносом протонов через мембраны, ведущим к образованию градиента электрохимических потенциалов ионов водорода (∆µН+). Целью настоящей работы являлось изучение влияния ИУК на параметры протонного переноса в хлоропластах и проводимости БЛМ, на процессы образования энергии в растительной клетке.
Материалы и методы исследования
Объектами исследования служили 2-20-дневные проростки растений, различающихся по радиочувствительности: конских бобов (Vicia faba), сои (Glicine hispida), кукурузы (Zea mays), подсолнечника (Helianthus annus), редис (Raphanus sativus). Облучение проводили на установке ГУПОС с источником излучения Cs137 при мощности дозы 500 р/мин. Семена облучали стимулирующими и угнетающими дозами, предварительно установленными для каждой культуры: для бобов - 100 р и 10 кр, для кукурузы - 500 р и 40 кр, сои - 700 р и 40 кр, подсолнечника - 1000 р и 64 кр, редиса - 1000 р и 300 кр. Семена высевали в отдельные сосуды с трехкратной повторностью, спустя 24 часа после облучения, и выращивали при освещении лампой БА-30.
Определение свободного триптофана проводили методом хроматографии на бумаге по Кудряшовой (1964) с модификацией. В качестве первого растворителя использовали смесь метанола с 0,01М раствором Na2HPO4 в соотношении 4:5. Вторым растворителем являлась дистиллированная вода. В этих условиях триптофан четко отделялся от остальных аминокислот в виде подковообразного пятна с Rf = 0,7-0,75. Этот метод позволяет также определять содержание суммы аминокислот - как свободных, так и связанных. Определение суммы свободного и связанного триптофана проводили по методу Смирновой-Иконниковой (1965) для сельскохозяйственных культур.
Активность триптофансинтазы определяли по методу Мадхузу-данана Наира и Вайдианатана (1961). Экстракцию фермента проводили раствором глутатиона в концентрации 10-3 М при to = 0-4°C в соотношении растительного материала (г) и раствора (мл) 1:4. Гомогенат центрифугировали 20 мин при 3000 g также при to = 0-4°C. Реакционную смесь для определения активности фермента инкубировали 3 часа при 25°С. Содержание триптофана, образовавшегося в результате действия триптофансинтазы, определяли колориметрически на ФЭК-М при красном светофильтре. Активность выражали в мкг образовавшегося триптофана за I час.
Выделение, очистку и количественное определение индольных производных проводили по разработанному нами методу [5], который позволил увеличить чувствительность и специфичность определений. Определяли содержание природных индольных производных, а также серина, суммы свободных и связанных аминокислот. Содержание белка определяли по Лоури.
Результаты обрабатывали статистически, достоверность данных оценивали для каждого варианта по критерию Стъюдента и по методу вычисления стандартной ошибки и доверительных интервалов средних арифметических величин.
Хлоропласты выделяли из листьев 12-14 дневных проростков гороха (Pisum sativum), выращенных в теплице. Полученная суспензия хлоропластов содержала пластиды типа В и С по классификации Хилла. Параметры протонного обмена измеряли на рН-метрической установке, собранной на базе ионометра ЭВ-74, самописца ТZ 215, термостатированной стеклянной ячейки и стандартных рН электродов. Среда реакции в ячейке объемом 1,8 мл содержала 50 мМ КСl, 10 мМ NаС1, 2 мМ МgCl2, количество хлоропластов соответствовало 50 мкг хлорофилла. Температуру реакционной среды поддерживали в пределах 20-21°С. Суспензию освещали красным светом (λ>600 нм) через светофильтр КС-13 с интенсивностью луча 100 Дж.м-2 с-1.
Модельные липидные мембраны БЛМ получали из общей фракции фосфолипидов мозга быка и сои с концентрацией 20 мг/мл в н-декане («Sigma», США). Электропроводность мембран измеряли по методике [11].
Результаты исследований
Влияние γ-облучения семян на биосинтез индольных производных в проростках было изучено при действии стимулирующих и угнетающих доз радиации на семена, влияние различных доз на рост мы сопоставили с накоплением и биосинтезом основного ауксина ИУК - гормона роста. Было показано, что под действием стимулирующих доз происходит стимуляция роста проростков растений, начиная с третьего дня развития и до конца опыта на 20,7-45,4% по отношению к контролю. Под влиянием угнетающих доз наблюдается резкое ингибирование их роста и затем гибель на ранних этапах развития (табл. 1).
Исследования влияния радиации в стимулирующих и угнетающих дозах на биосинтез ИУК проводили на 9-дневных проростках кукурузы. При воздействии γ-облучением в малых стимулирующих дозах (100р - 1кр) на радиочувствительные семена концентрация ИУК возрастает. При высоких угнетающих дозах радиации (10-60 кр) концентрация ИУК даже в радиоустойчивых семенах резко снижается, при этом происходит торможение роста, гибель проростков и образование мутаций. Результаты колориметрического определения индольных производных приводятся в табл. 2.
Таблица1
Влияние γ-радиации на динамику роста проростков кукурузы при γ-облучении семян
|
Доза |
3 день |
9 день |
14 день |
20 день |
||||
|
кр |
мм |
% |
мм |
% |
мм |
% |
мм |
% |
|
0 |
36,3±0,9 |
100 |
148,8±1,5 |
100 |
196,4±3,7 |
100 |
251,5±3,4 |
100 |
|
0,5 |
52,8±0,9 |
145 |
189,3±1,6 |
127 |
253,0±3,7 |
129 |
303,8±4,5 |
121 |
|
40 |
16,0±0,3 |
44 |
16,1±0,4 |
11 |
- |
- |
- |
- |
Таблица 2
Влияние γ-радиации на содержание индольных производных в 9-дневных проростках кукурузы (в мкг на 50 г абсолютного сухого веса)
|
Доза |
ИУК |
β-индолил-пировиноградная кислота |
β-индолил-3- пропионовая кислота |
|||
|
кр |
мкг |
% |
мкг |
% |
мкг |
% |
|
0 |
6,75±0,14 |
100 |
3,80±0,12 |
100 |
2,50±0,12 |
100 |
|
0,5 |
8,50±0,14 |
126 |
4,51±0,87 |
118,7 |
3,00±0,09 |
120 |
|
40 |
3,50±0,16 |
51,8 |
1,03±0,050 |
27,2 |
5,95±0,14 |
238 |
Из данных таблицы 2 следует, что количество ИУК, β-индолилпировиноградной кислоты и β-индолил-3-пропионовой кислоты под действием стимулирующих доз γ-облучения увеличивается по отношению к контролю на 26%, 18,7% и 20%, соответственно. Под действием угнетающих доз содержание ИУК уменьшается на 49,2%, β-индолилпировиноградной кислоты - на 72,8%, в то время как количество β-индолил-3-пропионовой кислоты увеличивается на 138%. Можно предположить, что синтез ИУК при угнетающих дозах нарушается на стадии образования β-индолилпировиноградной кислоты и происходит ингибирование активности специфичной дегидрогеназы, которая превращает β-индолил-3-пропионовую кислоту в β-индолилпировиноградную.
В настоящее время известно, что ИУК синтезируется также из триптофана, поэтому было предположено, что снижение содержания ИУК может происходить также за счет нарушения его синтеза, при этом первичные изменения при действии γ-облучения могут происходить на уровне кодирования генов в ДНК. Показано, что под влиянием стимулирующих доз γ-радиации содержание свободного триптофана в 9-20 дневных проростках всех исследуемых культур увеличивается на 13-22% (рис. 1). Облучение семян в угнетающих дозах вызывает уменьшение содержания свободного триптофана во всех культурах на 10-75%. Полученные данные подтверждают литературные, согласно которым при облучении семян ячменя в угнетающих дозах в 7-дневных проростках наблюдается уменьшение содержания триптофана на 50% [12].
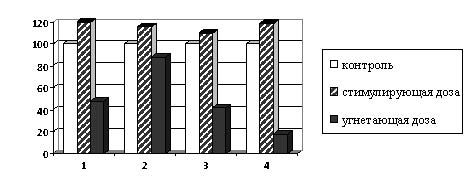
Рис. 1. Влияние γ-облучения на содержание триптофана в проростках
при предпосевном облучении семян (в % к контролю). 1 - конские бобы, 2 - соя,
3 - подсолнечник, 4 - редис
Так как триптофан в растениях может синтезироваться из серина и индола, мы провели исследования количественного изменения серина (рис.2).
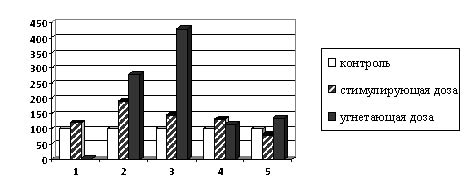
Рис. 2. Влияние γ-облучения на содержание серина в проростках
при предпосевном облучении семян (в % к контролю). 1 - конские бобы; 2 - соя;
3 - кукуруза; 4- подсолнечник; 5 - редис
Из данных, приведенных на рис. 2, следует, что под влиянием γ-облучения в стимулирующих дозах наблюдается тенденция к увеличению содержания свободного серина в проростках всех культур, что приводит к стимуляции синтеза триптофана из серина и индола. Под влиянием угнетающих доз радиации наблюдается увеличение количества серина: резкое - в радиочувствительных культурах (на 165% в сое и на 315% в кукурузе) и сравнительно небольшое увеличение его - в радиорезистентных. Такое различие, возможно, связано с тем, что у последних серин используется на синтез серосодержащих аминокислот - цистеина и метионина, обуславливающих радиорезистентность растений. Сопоставление полученных данных позволяет заключить, что при действии угнетающих доз радиации накопление серина связано с блокированием биосинтеза триптофана.
Интересно было провести определение активности фермента триптофансинтазы, катализирующего синтез триптофана из серина. Определение проводили в проростках растений при предпосевном облучении семян в стимулирующих и угнетающих дозах γ-облучения (табл. 3).
Таблица 3
Влияние γ-облучения на активность триптофансинтазы в проростках конских бобов, кукурузы и подсолнечника (мкг/час)
|
Доза облучения |
Дни прорастания |
|||
|
Конские бобы |
2 |
4 |
14 |
20 |
|
0 |
28,60±1,00 |
19,70±0,98 |
30,60±0,42 |
34,00±0,82 |
|
100 р |
32,46±1,20 |
25,60±0,95 |
35,30±0,65 |
36,40±0,94* |
|
10 кр |
26,46±0,37* |
14,06±0,42 |
22,60±0,71 |
26,40±0,34 |
|
Кукуруза |
6 |
9 |
14 |
- |
|
0 |
18,40±0,19 |
10,30±0,38 |
8,10±0,39 |
- |
|
500 р |
22,06±0,28 |
13,00±0,52 |
10,20±0,28 |
- |
|
40 кр |
15,70±0,15 |
6,90±0,31 |
6,30±0,23 |
- |
|
Подсолнечник |
4 |
9 |
14 |
20 |
|
0 |
17,10±0,22 |
16,00±0,32 |
20,60±0,30 |
13,60±0,35 |
|
I кр |
19,50±0,53 |
17,60±0,63* |
22,10±0,58* |
15,00±0,75* |
|
64 кр |
13,06±0,48 |
12,00±0,62 |
14,26±0,47 |
9,00±0,11 |
*) Разница от контроля недостоверна.
Из таблицы 3 видно, что при облучении семян стимулирующими дозами радиации активность фермента в проростках всех изученных нами культур увеличивается (в конских бобах - на 7-30%, кукурузе - на 20-26%, подсолнечнике - на 7-14%), что не противоречит небольшому увеличению содержания серина - субстрата данного фермента. При облучении высокими дозами - активность фермента снижается (в конских бобах - на 8-29%, в кукурузе - на 15-33%, в подсолнечнике - на 24-34%), что согласуется с существенным накоплением серина. При облучении высокими дозами облучения тех же видов семян на Серпуховском ускорителе и малыми дозами γ-облучения на установке ИБК РАН были получены аналогичные данные по активации и угнетению активности фермента.
Нами было изучено действие ИУК на протонную проводимость тилакоидных биологических мембран. На рис. 3а видно, что с увеличением концентрации ИУК величина стационарного протонного градиента [Н+]ст. уменьшается в диапазоне рН 6,0-7,0, что коррелирует с увеличением пассивной протонной проницаемости (КТ) мембраны (рис. 3б) в этом же диапазоне рН.
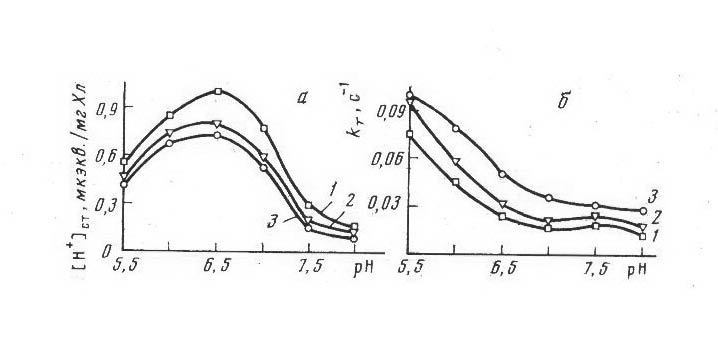
Рис. 3. Зависимость величины стационарного протонного поглощения (а) и пассивной протонной проницаемости (б) мембраны тилакоидов от концентрации ИУК:
1 - контроль, 2 - 5 . 10-7 М, 3 - 1 . 10-4 М
Выяснено, однако, что основное (до 15%) уменьшение [Н+]ст. происходит при низких концентрациях ИУК (5.10-7 М), тогда как пассивная проницаемость при той же концентрации увеличивается незначительно и резко возрастает (до 100%) при 10-4 М. Приведенные результаты по действию ИУК на тилакоидные мембраны показывают, что эффект ауксина более заметен при слабокислых и нейтральных значениях рН. В оптимальных условиях реакции фотовосстановления эффект ИУК незначителен как при малых, так и при больших концентрациях вещества.
Была измерена также величина КТ [Н+]СТ. (рис. 4а), которая равна скорости выхода протонов из тилакоидов в момент выключения света, она обычно принимается равной скорости выхода протонов на свету [18]. В стационарных условиях освещения скорость выхода протонов равна скорости поступления их в тилакоиды, которая непосредственно связана со скоростью транспорта электронов. Таким образом, увеличение КТ [Н+]СТ. свидетельствует об увеличении скорости электронного транспорта при увеличении концентрации ИУК.
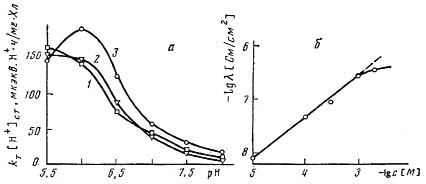
Рис. 4. Зависимости: а) скорости поступления протонов внутрь тилакоидов при постоянном освещении от рН при различных значениях концентрации ИУК
(1-3 - см. рис. 3); б) протонной проводимости БЛМ от концентрации ИУК.
Согласно современным представлениям, одним из структурных компонентов биологической мембраны является липидный бислой, служащий основным барьером для транспорта вещества. Следовательно, ионофор, транспортирующий ионы через липидный бислой, будет переносить их и через биологические мембраны [11]. Ранее нами было выяснено, что под действием многих слабых кислот [11,13,14] может происходить увеличение проницаемости БЛМ для протонов, что проявляется в увеличении их электропроводности. Поскольку ИУК также является слабой кислотой с рК = 4,7, можно было ожидать, что она также будет увеличивать проницаемость БЛМ. Нами была изучена зависимость проводимости БЛМ от концентрации ИУК в буферной среде (20 мМ трис-НСI, рН 7,5) со значением рН, близким к концентрации водородных ионов большинства физиологических сред. При увеличении концентрации ИУК от 10-5 М до 10-4 М в растворе проводимость (-lgс [М]) мембран увеличивается в 5 раз, при дальнейшем увеличении концентрации ауксина до 10-3 М проводимость возрастает еще на порядок с последующим выходом на плато. На БЛМ из соевых растительных липидов проводимость мембран была выше на порядок при тех же концентрациях. При высоких концентрациях происходит насыщение ИУК в самой мембране, либо в пограничном слое. Из линейной зависимости проводимости БЛМ от концентрации вещества можно предположить, что в переносе одного иона Н+ через мембрану участвует одна молекула модификатора. Возможно, что лимитирующей стадией переноса протона является химическая реакция между модификатором и переносимым ионом на границе мембраны.
Обсуждение результатов
Показано, что облучение семян низкими, стимулирующими рост и развитие проростков дозами γ-радиации приводит к увеличению концентрации ИУК, а высокими, ингибирующими дозами - к ее уменьшению. Выявлены радиочувствительные и радиоустойчивые виды семян. Содержание ИУК изменяется под действием γ-облучения за счет изменения скорости ее образования двумя путями. Первый путь синтеза ИУК - из триптофана. При γ-облучении семян в стимулирующих дозах увеличение концентрации ИУК согласуется с незначительным увеличением содержания в проростках свободного триптофана и увеличением активности соответствующего фермента триптофансинтазы. При γ-облучении в угнетающих дозах содержание триптофана существенно уменьшалось в результате снижения скорости синтеза из серина и индола (угнетается активность триптофансинтазы, значительно увеличивается содержание серина). Второй путь биосинтеза ИУК - из β-индолилпировиноградной кислоты и триптамина. Под действием стимулирующих доз γ-облучения он также активируется, а угнетающих - ингибируется, - на стадии образования β-индолилпировиноградной кислоты из β-индолил-3-пропионовой. Действие стимулирующих доз радиации приводило к усилению белкового синтеза и связанному с ним уменьшению содержания суммы свободных аминокислот, угнетающих доз - к увеличению их содержания, что можно объяснить усилением действия протеаз и гидролитических процессов, возникающих в растении, облученном высокими дозами γ-радиации.
Нами были изучены также механизмы действия ИУК на рост и развитие растений. ИУК является ауксином - растительным гормоном, обладающим высокой физиологической активностью в очень низких концентрациях (пико- и нанограммы на г сырой биомассы) [8]. В литературе приводятся данные о взаимодействии ИУК и других фитогормонов (цитокининов) с рецепторами, которые локализованы в плазмалемме, эндоплазматической сети и цитоплазме растягивающихся клеток. Одним из результатов этого взаимодействия является активация Н+-помпы, Н+, К+-АТФазы плазмалеммы и выделение ионов Н+ из клеток, одновременно развивается гиперполяризация мембранного потенциала цитоплазматической мембраны. Выброс Н+ происходит в обмен на ионы Са2+ и К+, при увеличении концентрации Са2+ до 10 мМ увеличивается влияние ИУК на возникший градиент биоэлектропотенциалов (БЭП) [9]. БЭП является потенциалом действия, в данном случае роста, и индуцируется, как правило, при сильных воздействиях факторов внешней и внутренней среды. Отличаясь по величине в различных частях растения, БЭП может распространяться из клетки в клетку по плазматической мембране со скоростью 0,1-1,0 см/сек в виде медленных волн с периодом 1-10 мин. ИУК в комплексе с рецептором достигает ядра и активирует в нем синтез РНК, а затем синтез аминокислот и белков. ИУК, таким образом, влияет на генную активность и проницаемость цитоплазматических мембран. Энергетические затраты на эти процессы обеспечиваются усилением дыхания [8].
При использовании низких физиологических концентраций ИУК на мембранах тилакоидов нами было показано снижение величины [Н+]ст. и увеличение пассивного выхода Н+. Было показано, что ИУК обладает слабым протонофорным действием при низких концентрациях и увеличивает пассивную проницаемость тилакоидной мембраны для ионов водорода, направленную из тилакоидов в цитоплазму клетки. Мы полагаем, что стимуляция пассивной Н+ проницаемости тилакоидов обусловлена подщелачиванием цитоплазмы при активации ауксином цитоплазматической Н+-АТФазы, осуществляющей выброс протонов из клетки. Подтверждением протонофорности служат наши данные, полученные на БЛМ, и литературные на плоских липидных мембранах [15]. Наблюдаемое нами слабое, временное разобщение между образованием энергии и переносом протонов по ЭТЦ тилакоидов в условиях in vivo может приводить к последующему увеличению градиента протонов на их мембране и биосинтеза АТФ в процессе фотофосфорилирования (ФФ) в тилакоидах. Аналогичные данные были получены нами при разобщении окислительного фосфорилирования в митохондриях млекопитающих [11]. В соответствии с нашим предположением, при фосфорилирующих условиях на изолированных хлоропластах гороха было показано, что ИУК в малых концентрациях (10-10 - 10-7 М) ускоряла эндогенный тип ФФ на 80%, синтез АТФ с феназинметасульфатом - на 50%, физиологический тип ФФ с НАДФ+ и ферредоксином - на 25% [1,9]. Полученные авторами результаты зависели от того, какие в опытах использовались акцепторы электронов - искусственные или природные. Полагают, что ИУК стимулирует фотосинтетический электронный транспорт, оказывая ингибируюшее действие на участок восстановления НАДФ+, что возможный механизм действия гетероауксина и других фитогормонов связан с регуляцией ферментных систем, участвующих в образовании НАДФН и АТФ [6]. На основании представленных нами данных обосновывается еще один, энергетический путь влияния ИУК на рост растений, наряду с известными путями - гормональным и генетическим. Нами показано, что высокие нефизиологические концентрации ИУК (выше 10-4 М) вызывают значительное увеличение протонной проводимости БЛМ и сильное разобщение ФФ на хлоропластах, что согласуется с данными зарубежных авторов [16].
Показано, что при использовании γ-облучения или адронного излучения в разных дозах возможно увеличить или снизить концентрацию ИУК в проростках из облученных семян, незначительные ее изменения достаточны для получения возможности регулировать рост и урожайность сельскохозяйственных культур.
СПИСОК ЛИТЕРАТУРЫ
- Акулова Е.А., Мурзаева С.В. и др. // Биохимия. 1975. Т. 40. Вып. 6. С. 1205.
- Березина Н.М. // Предпосевное облучение семян сельскохозяйственных культур γ-лучами. 1969. Пущино.
- Кузин А.М. // Стимулирующее действие ионизирующего излучения на биологические процессы. 1977. М.: Атомиздат. 136 с.
- Ревин А.Ф. // Физиол. растений. 1970. Вып. 6. С. 1271.
- Ревин А.Ф., Смолянинов В.В. // Способ очистки индольных производных производных класса гетероауксинов в растительном сырье. Авт. свид. № 362830. 1973. Бюлл. № 3.
- Рощина В.Д., Боровикова Л.В., Рощина В.В., Шерудило Е.Г. // Физиол. растений. 1980. Т. 27. Вып. 6. С. 1267.
- Полевой В.В. // Фитогормоны. 1982. Л.: ЛГУ. 249 с.
- Полевой В.В. // Соросовский образовательный журнал. 1997. № 9. С. 6.
- Музафаров Е.Н., Креславский В.Д., Назарова Г.Н. // Световая и гормональная регуляция фотосинтеза и роста растений. 1995. Пущино. ОНТИ ПНЦ РАН. 140с.
- Неминущий В.Г., Каюшин Л.П., Кожокару А.Ф., Назарова Г.Н., Музафаров Е.Н. // Биофизика. 1987. Вып. 32. № 2. С. 352.
- Кожокару А.Ф. // «Направленная модификация структурно-функционального состояния мембран с целью изменения устойчивости клеток и организма к факторам окружающей среды». 1992. Дис. ... д.ф-м.н. М. МНО «Форум». 88 с.
- Хвойка Л., Кутачек М. и др. // Использование изотопов и излучений в исследованиях по сельскому хозяйству. 1967. М.: Колос.
- Cojocaru A.F., Ruzieva R.H. et al. // Studia biophysica. 1978. V.72. № 1. P. 15-30.
- Cojocaru A.F., Muzafarov E.N. et al.//Studia biophysica.1988.V.126. № 3. P.141-156.
- Guthnecht I., Walter A.J. // J. Membr. Biol. 1980. V. 56. № 1. P. 65.
- Driessche T.V., Lannoye R. // Int. J. Chronobiol. 1982. V. 8. № 2. Р. 97.
- Мitchell P. // Chemiosmotic coupling in oxidative and photosynthetic phosphorylation. 1966. Bodmin-Cornwall: Glynn Research.
- Schawartz M. // Ann. Rev. Plant Physiol. 1971. V. 22. P. 469.
Библиографическая ссылка
Кожокару А.Ф., Ревин А.Ф. ИЗУЧЕНИЕ МЕХАНИЗМА ДЕЙСТВИЯ γ-ОБЛУЧЕНИЯ НА БИОСИНТЕЗ ИНДОЛЬНЫХ ПРОИЗВОДНЫХ И ИХ РОЛИ В ОБРАЗОВАНИИ ЭНЕРГИИ В ПРОРОСТКАХ РАСТЕНИЙ // Современные проблемы науки и образования. – 2010. – № 4. ;URL: https://science-education.ru/ru/article/view?id=4493 (дата обращения: 23.04.2024).