



Активно проводимые исследования патогенеза боли как у нас в стране, так и за рубежом, позволили сформировать достаточно полное представление о триггерных механизмах боли, морфологическом и биохимическом субстратах боли, ее особенностях при различных заболеваниях и принципах патогенетической терапии. Налицо концентрация интересов исследователей на «ядре» боли, ее «веерные» механизмы удостоены несравненно меньшего внимания.
В литературе крайне ограничена информация о роли врожденного иммунитета (неспецифической резистентности) организма в механизмах боли [1]. Большинство трудов, написанных видными учеными, посвящено изучению специфических иммунологических ответов на системном, органном, клеточном и субклеточном уровнях, а фагоцитозу в них отводится лишь справочная роль. Однако более целесообразно начинать изучение механизмов защиты с фагоцитоза как наиболее ранней, срочной и эффективной неспецифической реакции организма [3].
Цель исследования
Очевидно, что механизмы фагоцитоза, его регуляция мало изучены и приобретают самостоятельное значение в контексте патогенеза боли. Это направление, преломленное через призму возрастной патологии, практически не изучено, что и определяет актуальность проблемы и интерес к ней. Поэтому наше исследование посвящено проблеме изучения онтогенетических аспектов изменения фагоцитарной активности лейкоцитов при острой соматической боли.
Материал и методы исследования
Исследования выполнены на 180 неполовозрелых нелинейных белых крысах обоего пола без учета половых различий. Все экспериментальные животные были разделены на 2 возрастные группы: новорожденные (3-4 дневные) - 90 крыс, и месячные - 90 крыс. В работе выполнено 3 серии исследований: 1 серия - определение фагоцитарной активности (фагоцитарный индекс (ФИ) и фагоцитарное число (ФЧ)), 2 серия - изучение постболевых изменений компонентов цитохимических реакций в лейкоцитах (пероксидаза, щелочная фосфатаза и гликоген), 3 серия - изучение изменений внутрисистемных гуморальных регуляторов белой крови - интерлейкинов (ИЛ) (ИЛ - 1α, ИЛ - 4, ИЛ - 6, фактор некроза опухоли (ФНОα)). В каждой возрастной группе исследовали исходный фон, изменения через 2 минуты и через 60 минут после острого болевого воздействия (ОБВ).
Острую соматическую боль (ОСБ) 4-5 степени интенсивности [5] моделировали путем электрокожного раздражения рецепторной зоны корня хвоста крыс с помощью электростимулятора ЭСУ-2.
Забор материала для исследования производили на пике развития ноцицептивной реакции (через 2 минуты после электростимуляции) и в посттравматическом периоде (через 60 минут после электростимуляции). Все манипуляции с экспериментальными животными проводили в соответствии с Приказом МЗ РФ №267 «Об утверждении правил лабораторной практики» от 19 июня 2003 г. Для определения фагоцитарной активности лейкоцитов использовали метод Д. К. Новикова in vitro [4]. Для определения пероксидазы в лейкоцитах использовали метод Леле (реакция с бензидином) [8]. Для определения щелочной фосфатазы в лейкоцитах использовали метод азосочетания по Хейхоу и Кваглино [9]. Для исследования содержания гликогена в лейкоцитах использовали метод ШИК-реакции по Мак-Манусу [6]. Концентрацию интерлейкинов (Il-1α, Il-4, Il-6) и ФНО-α в сыворотке крови экспериментальных крыс определяли методом иммуноферментного анализа с помощью наборов фирмы Bender MedSystems (Europe). Приготовление растворов стандартов, конъюгатов, промывочного и рабочего буфера проводили согласно инструкции к набору [10]. Достоверность различий по количественным признакам определяли с помощью критерия Стьюдента (t) для малых выборок, признавая их статистически значимыми при p≤0,05, как тенденцию к достоверности при 0,05<p≤0,1. Достоверность различий качественных показателей при сравнении оценивали с помощью непараметрического критерия статистики (χ) ван дер Вардена [7].
Результаты исследования. Обсуждение
Через 2 минуты после нанесения болевого раздражения фагоцитарный индекс не имеет статистически существенного отличия от зарегистрированного в исходном состоянии, но в то же время фагоцитарное число увеличивается. За 60 минут с начала болевой стимуляции показатели, характеризующие фагоцитарную активность, практически не претерпевают изменений. Создается впечатление, что сильное болевое воздействие исчерпало все резервы активации фагоцитарной активности сразу, и они не имеют в динамике процесса поступательного развития. Так, при незначительном увеличении фагоцитарного индекса фагоцитарное число практически не меняется (табл. 1).
Табл. №1
|
Изменения фагоцитарной активности у новорожденных крыс |
|||
|
Группы |
ФИ, % |
ФЧ |
|
|
Исходный фон |
50,4±1,8 |
2,1±0,1 |
|
|
ОБВ |
через 2 мин |
47,0±0,6 |
2,7±0,11 |
|
через 60 мин |
49,9±0,92 |
2,6±0,11 |
|
1 - различия достоверны по отношению к исходному фону, p≤0,05.
2 - различия достоверны между группами с острым болевым воздействием, p≤0,05 (по t - критерию Стьюдента).
Через 2 минуты после болевого воздействия у крыс месячного возраста фагоцитарный индекс и фагоцитарное число снижается по сравнению с исходным состоянием. Через 60 минут фагоцитарное число продолжает оставаться сниженным в сравнении с исходным состоянием, однако, сами фагоциты постепенно восстанавливают активность: фагоцитарное число стремится приблизиться к исходным значениям (табл. 3).
Табл. №3
|
Изменения фагоцитарной активности у крыс месячного возраста |
|||
|
Группы |
ФИ, % |
ФЧ |
|
|
Исходный фон |
53,2±1,2 |
3,1±0,1 |
|
|
ОБВ |
через 2 мин |
42,8±11 |
2,1±0,112 |
|
через 60 мин |
40,3±0,81 |
2,8±0,1 |
|
1 - различия достоверны по отношению к исходному фону, p≤0,05.
2 - различия достоверны между группами с острым болевым воздействием, p≤0,05 (по t - критерию Стьюдента).
Щелочная фосфатаза обеспечивает микробицидные свойства нейтрофильных лейкоцитов, подготавливает микроорганизмы к их дальнейшей дезинтеграции пероксидазой. Пероксидаза катализирует окисление хлорид аниона перекисью водорода и вместе с ним образуют мощный бактерицидный агент - гипохлорит [2].
Активность фагоцитоза, его завершенность зависят от уровня гликогена, основного депо энергетического обеспечения фагоцитирующей клетки. Об изменениях, изучаемых, цитохимических компонентов свидетельствует интегральная величина, характеризующая динамику биохимических процессов - средний цитохимический коэффициент (К).
Активность щелочной фосфатазы как у новорожденных, так и у особей препубертатного периода примерно одинакова Активность пероксидазы выше у крыс препубертатного возраста. В структуре распределения нейтрофилов по степени их насыщенности пероксидазой у новорожденных присутствуют только нейтрофилы, содержащие малое и умеренное количество гранул. В отличие от новорожденных, у крыс месячного возраста клетки с максимальной насыщенностью пероксидазой присутствуют в 16 %. Распределение гликогена в лейкоцитах крыс разного возраста не имеет существенных различий (табл.4, табл.5).
Табл. №4

1 - разница существенна, в сравнении с исходным фоном, на уровне доверительной вероятности 0,05 (5%) (по Х критерию ван дер Вардена).
2 - различия достоверны по отношению к исходному фону, p≤0,05 (по t - критерию Стьюдента).
3 - различия достоверны между группами с острым болевым воздействием, p≤0,05 (по t - критерию Стьюдента).
Таким образом, распределение щелочной фосфатазы и гликогена в нейтрофилах интактных новорожденных животных и животных препубертатного возраста практически тождественно, активность пероксидазы выше у крыс месячного возраста, что можно расценивать как более высокую готовность к отражению микробной агрессии. С возрастом происходит активация (созревание) кислородзависимых механизмов фагоцитоза - системы «респираторного взрыва», одним из компонентов которой является пероксидаза. Так как «респираторный взрыв» базируется на пентозофосфатном окислении глюкозы, становится понятной тенденция возрастного снижения содержания гликогена - энергетическое депо.
Проведенный анализ показал, что ответная реакция на боль у животных месячного возраста отличается от таковой у новорожденных (табл.5).
Табл. №5
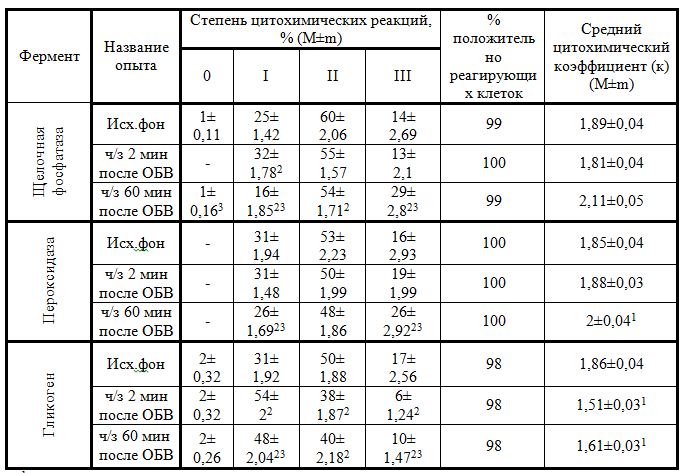
1 - разница существенна, в сравнении с исходным фоном, на уровне доверительной вероятности 0,05 (5%) (по Х критерию ван дер Вардена).
2 - различия достоверны по отношению к исходному фону, p≤0,05 (по t - критерию Стьюдента).
3 - различия достоверны между группами с острым болевым воздействием, p≤0,05 (по t - критерию Стьюдента).
В ней нет выраженной однонаправленности, она носит фазный характер. Через 2 мин после интенсивного болевого воздействия средний цитохимический коэффициент снижается. Через 60 мин баланс смещается, цитохимический коэффициент имеет тенденцию к увеличению (р≤0,1).
Проведенное исследование убедительно показало, что активность пероксидазы лейкоцитов новорожденных крыс практически не меняется в ответ на нанесение болевого раздражения ни сразу, ни после временной экспозиции. Крысы препубертатного возраста в отличие от новорожденных дают достаточно выраженную цитохимическую реакцию. Активность пероксидазы динамична и дифференцирована в зависимости от времени эксперимента. Общая направленность изменения показателей свидетельствует о нарастании активности фермента после электростимуляции, что свидетельствует о функциональном созревании кислородзависимых механизмов фагоцитоза. Гликоген нейтрофилов новорожденных крыс лабилен, оперативно включается в реакцию на болевое раздражение. Наблюдается уверенное и последовательное снижение присутствия гранул гликогена в нейтрофильных лейкоцитах. Обмен гликогена в цитоплазме нейтрофильных лейкоцитов у животных месячного возраста более обстоятельный, рациональный и адекватный, нежели у новорожденных крыс. Для него характерна двухфазность. Первичная реакция на боль ведет к потере гранул, но уже через час содержание гликогена обретает явную тенденцию к восстановлению.
Полученные факты позволяют заключить, что у новорожденных животных более активна щелочная фосфатаза, а у месячных - пероксидаза, т.е. в периоде новорожденности преобладают кислороднезависимые механизмы бактериоцидности, а в месячном возрасте - кислородзависимые. Истощение активности гликогена в большей степени отмечается у новорожденных животных, оно нарастает в динамике эксперимента. Очевидно, активация фагоцитоза у них более энергозатратна.
Результаты исследований цитокинов представлены в таблице 6.
Табл.6 Исследования изменения уровня цитокинов в сыворотке крови при ОСБ у крыс в онтогенезе
|
Возраст |
Категория |
Цитокины |
|||
|
ИЛ 1 |
ИЛ 4 |
ИЛ 6 |
ФНОα |
||
|
Новорожденные |
исходный фон |
4,85±0,29 |
2,67±0,32 |
36,55±4,12 |
60,37±6,65 |
|
2 мин после ОБВ |
6,6±0,51 |
6,71±0,431 |
58,41±4,671 |
38,08±3,021 |
|
|
60 мин после ОБВ |
5,68±0,6 |
4,14±0,3812 |
41,74±5,512 |
30,25±2,6812 |
|
|
Месячные |
исходный фон |
6,49±0,53 |
3,31±0,41 |
50,95±6,53 |
18,35±1,77 |
|
2 мин после ОБВ |
5,95±0,69 |
7,8±0,471 |
56,08±4,58 |
21,66±3,99 |
|
|
60 мин после ОБВ |
8,54±0,4112 |
4,38±0,512 |
49,59±13,33 |
28,72±2,121 |
|
1 - различия достоверны по отношению к исходному фону, p≤0,05.
2 - различия достоверны между группами с острым болевым воздействием, p≤0,05 (по t - критерию Стьюдента).
В сыворотке крови новорожденных крыс после ОБВ уже через 2 минуты отмечается увеличение уровня ИЛ-1α. Новорожденные животные отвечают двухфазной реакцией: срочным подъемом уровня ИЛ-1α с последующей тенденцией к нормализации к концу первого часа, у крыс месячного возраста в течение часа реакция имеет однонаправленный характер, отсутствие ответа в первые две минуты эксперимента и явным увеличением к концу первого часа.
У новорожденных через 2 минуты в содержание ИЛ-6 регистрируется статистически подтвержденный количественный всплеск интерлейкина. Животные препубертатного возраста имели фоновые значения ИЛ-6 в полтора раза выше новорожденных. Таким образом, у новорожденных животных отмечается резкий подъем ИЛ-6 через 2 минуты после ОБВ, с практически полным возвратом показателей к исходным, к концу первого часа, а его содержание у крыс препубертатного возраста сохраняется примерно на одном уровне в динамике всего эксперимента.
Наиболее определяемым в количественном отношении явился ФНО-α. Нанесение ОБВ новорожденным животным вызвало реакцию, полярную той, которая отмечалась в отношении ИЛ-1α и ИЛ-6. Регистрируется уменьшение уровня ФНО-α по сравнению с исходным. Через час содержание ФНО-α у новорожденных продолжало снижаться. В исходном состоянии содержание ФНО-α у месячных крыс втрое ниже, чем у новорожденных. Нанесение ОБВ увеличивает содержание исследуемого цитокина, хотя различие между рядами статистически не существенно. Через час после ОБВ количество ФНО-α увеличивается (р≤0,01).
После ОБВ содержание ИЛ-4 возрастает. Через 1 час в сыворотке остается увеличенным содержание ИЛ-4. Аналогичная по фазности развития картина наблюдалась и у крыс препубертатного возраста. Через 2 минуты после ОБВ зарегистрирован достаточно выраженный подъем интересующего нас цитокина. В течение часа прослеживается снижение ИЛ-4, его количество приближается к исходному уровню. Таким образом, у животных обеих возрастных групп содержание ИЛ-4 нарастает вслед за ОБВ уже через 2 минуты, спустя час после ОБВ ИЛ 4 стремится к своим исходным значениям.
Заключение
На основе литературного анализа наших данных можно сделать следующие заключения.
Одной из систем, закономерно реагирующих на боль, является система неспецифической фагоцитарной активности лейкоцитов. Степень реагирования врожденного иммунитета на боль зависит от возраста животных.
ОБВ приводит к активации ферментов, обеспечивающих микробицидность. Однако насколько интенсивна реакция, настолько она и истощаема, прежде всего, за счет ее энергозатратности. Бурный ответ, по всей видимости, сдерживается особенностями интерлейкинового обеспечения фагоцитарной активности, в частности, снижением ФНО-α.
У животных препубертатного возраста реакция лейкоцитов на боль более координирована. Она менее бурная, прослеживаются синхронная активация ферментов, обеспечивает переваривающую функцию фагоцитов, однако, и в этой группе животных энергетический потенциал нейтрофилов быстро исчерпывается.
В целом у животных раннего возраста фагоцитарная система функционально еще не готова к обеспечению полноценной неспецифической резистентности.
Рецензенты:
- Шестопалов А. В., д.м.н., зав. кафедрой общей и клинической биохимии №2,ГБОУ ВПО РостГМУ Минздравсоцразвития, г. Ростов-на-Дону.
- Харсеева Г. Н., д.м.н., профессор, зав. кафедрой микробиологии №2, ГБОУ ВПО РостГМУ Минздравсоцразвития, г. Ростов-на-Дону.
Библиографическая ссылка
Овсянников В.Г., Бойченко А.Е., Николаев В.Е., Бордюшков Ю.Н., Алексеев В.В., Алексеева Н.С. ОНТОГЕНЕТИЧЕСКИЕ ОСОБЕННОСТИ ИЗМЕНЕНИЯ ФАГОЦИТАРНОЙ АКТИВНОСТИ ЛЕЙКОЦИТОВ ПРИ ОСТРОЙ БОЛИ // Современные проблемы науки и образования. – 2012. – № 2. ;URL: https://science-education.ru/ru/article/view?id=5846 (дата обращения: 16.04.2024).