



Введение
Одним из основных последствий антропогенного воздействия на таежные экосистемы является уничтожение зональных сообществ при сведении лесов. Для выявления путей их эффективного возобновления необходимо изучение сохранившихся коренных лесов, не подверженных прямому влиянию человека. Важная роль в поддержании функционирования экосистем принадлежит почвенным сообществам. Адекватным индикатором состояния почвенно-растительного покрова являются насекомые, а в качестве биоиндикатора часто используются жужелицы [7, 12, 16, 33, и др.].
В многочисленных публикациях прослеживаются взаимосвязь факторов окружающей среды и состояния населения жужелиц в сообществах хвойно-широколиственных и южнотаежных лесов [9, 10, 11, 21, 24, 27, 29, и др.]. В то время как для жужелиц средней и северной тайги таких работ пока немного [3, 4, 13, 30].
Недостаточно также изучены жужелицы Вологодской области с фаунистической точки зрения. Имеется несколько работ, в которых содержатся сведения об их видовом разнообразии в лесных и околоводных сообществах в юго-западной и в центральной частях области [4, 24, 25]. В же время фауна региона представляет интерес, поскольку территория области находится в переходной зоне от полярных к умеренным широтам. Здесь проходила граница последнего оледенения, что повлияло на формирование биоты. На территории области сформировались леса двух подзон тайги южной и средней; сохранились элементы широколиственных и смешанных лесов. К тому же, в послеледниковое время территория области заселялась с запада и востока, что привело к взаимопроникновению западных (европейских) и восточных (сибирских) форм [5].
Целью настоящей работы является изучение структуры комплексов жужелиц коренных мало нарушенных среднетаежных лесов Вологодской области и выделение факторов, влияющих на их формирование.
Материалы и методы исследования
Исследования проводились в Шалго-Бодуновском ландшафтном заказнике в центральной части Вологодской области. Массив располагается в заповедной зоне национального парка «Русский Север», в пределах которой запрещена любая хозяйственная деятельность. Это позволяет рассматривать Шалго-Бодуновский заказник как эталонный среднетаежный лесной массив не подвергающийся прямому антропогенному воздействию. Кроме того, для него характерно значительное биотопическое разнообразие, что определяется сложным пересеченным рельефом территории [23]. Это также послужило причиной выбора района исследования. Сбор жуков осуществлялся в нескольких наиболее типичных для района исследования биотопах на различных элементах мезорельефа. Часть площадок были расположены вдоль линии профиля от вершины холма к межхолменному понижению и граничили друг с другом. Это сосняк-ельник чернично-зеленомошный на вершине холма и ельник папоротниково-таволговый в межхолменном понижении, а также соответственно ельник чернично-кислично-зеленомошный и ельник хвощово-папоротниково-кисличный. Остальные площадки (ельник болотно-травяный, ельник хвощово-папоротниковый, сосняк морошково-чернично-сфагновый) были расположены в межхолменных понижениях и не имели общих границ.
Исследования проводились с мая по октябрь в 2004 и 2005 годах. Для сбора насекомых применялась стандартная методика с использованием ловчих банок Барбера. Всего было установлено 150 ловушек. Собрано и обработано 1100 экземпляров имаго жужелиц.
Результаты
В ходе исследования на территории Шалго-Бодуновского заказника было обнаружено 27 видов жужелиц из 13 родов. Подобный уровень видового богатства характерен для таежных лесов. Так в южнотаежных хвойных лесах отмечено 22 - 32 вида жужелиц [9, 24]. Для лесных сообществ средней тайги указывается 26 - 36 видов [11, 19, 26], а для хвойных и хвойно-мелколиственных лесов северной тайги - 29 видов [30]. В то же время, общее количество видов на территории исследованного массива меньше числа видов, известных для лесных биоценозов средней тайги (49 видов) [30].
В составе фауны Шалго-Бодуновского леса преобладают виды с широким географическим распространением. Так, с позиций долготной составляющей (ареалы видов указываются по А. Г. Воронину [6]) преобладают европейско-сибирские (8) и трансевразийские (8) виды. Меньше видов с европейско-западносибирским (6) и голарктическим распространением (5).
Наибольшим видовым богатством в составе фауны жужелиц характеризуются рода Pterostichus (9 видов), Agonum (3 вида). Остальные рода включают по 1-2 вида. Сходный состав преобладающих родов отмечен для лесных сообществ и другими авторами [10]. Основу комплексов жужелиц на территории Шалго-Бодуновского массива составляют широко распространенные и многочисленные в лесной зоне виды [8, 13, 19, 27, 29, 30 и др.]: Pterostichus oblongopunctatus F. (23,17%), Calathus micropterus Duft. (18,13%), Epaphius secalis Pk. (12,82%), Carabus glabratus Pk. (7,78%). Значительный вклад в население жужелиц вносят такие виды как Agonum fuliginosus Pz. (4,12%), Patrobus assimilis Pk. (4,30%), Pterostichus niger Shall. (4,49%). Редкими являются 6 видов, зарегистрированные по 1-2 экземплярам: Notiophilus aquaticus (L.), Badister bullatus Schrank, Pterostichus quadrifoveolatus Letzn., Agonum gracile Gyll., A. micans Nic., Platynus mannerheimii Dej. Большинство из перечисленных видов довольно обычны для лесных или лесо-болотных сообществ тайги и в частности Вологодской области. В фаунистическом отношении интересны находки вида Platynus mannerheimii Dej., который распространен по всей зоне тайги, но почти везде является редким [1, 18, 30, 35]. А также вида Pterostichus quadrifoveolatus Letzn., который по литературным данным встречается спорадически и приурочен преимущественно к широколиственным лесам и южной тайге [1, 6, 15, 16, 31].
Изученные комплексы жужелиц лесных биотопов различаются по числу видов. Наиболее высокие показатели видового богатства характерны для влажных еловых лесов в межхолменных понижениях (18 - 8 видов). Также высокие показатели видового богатства комплекса жужелиц отмечены для ельника чернично-кислично-зеленомошного (16 видов), расположенного на вершине холма. Минимальное число видов жужелиц (по 7) входит в состав комплекса сосняка морошково-чернично-сфагнового и сосняка-ельника чернично-зеленомошного. Полученные данные соотносятся со сведениями других авторов о видовом богатстве жужелиц в таежных хвойных лесах. Так для южнотаежных ельников-кисличников указывается от 22 до 25 видов [9, 24], хвойных среднетаежных лесов 10 - 16 [11, 19], а хвойных массивов северной тайги - 12 - 13 видов [30].
Комплексы жужелиц биотопов Шалго-Бодуновского леса различаются по показателям суммарной уловистости. Были отмечены колебания данного параметра от 0,1 до 1,28 экз./10 ловушко-суток (далее экз./10 лов.-сут.). Уловистость жужелиц в изученных биотопах меньше аналогичных показателей для сообществ хвойных лесов южной и средней тайги. Так для комплексов жужелиц южнотаежных ельников отмечена уловистость 6,21 - 10,81 экз./10 лов.-сут., а для сосняков зеленомошников - 5,36 экз./10 лов.-сут. [9, 24]. Уловистость жужелиц в елово-черничных среднетаежных лесах составляет 1,65 экз./10 лов.-сут. [11]. Полученные в ходе исследований показатели уловистости близки к таковым для жужелиц северной тайги. Так, в северотаежных хвойных чернично-зеленомошных, еловых травяно-кустарничковых и сосновых чернично-сфагновых лесах показатели уловистости жужелиц изменяются от 0,31 до 0,96. экз./10 лов.-сут. [30].
Следует отметить, что показатели уловистости жужелиц в изученных биотопах увеличиваются по мере роста видового богатства их комплексов. Так при увеличении числа видов от 7 до 16 уловистость также возрастает от 0,1 до 1,28 экз./10 лов.-сут. Однако в комплексах с большим числом видов жужелиц (17 и 18 видов) происходит снижение уловистости жуков до 0,73 - 0,45 экз./10 лов.-сут.
Большинство жужелиц (13 видов) имеют широкое распространение в массиве и встречаются в различных типах биотопов на разных элементах мезорельефа. Однако есть виды, обитающие только в одном местообитании или нескольких биотопах сходных по условиям. Среди жужелиц, имеющих ограниченное распространение в массиве, преобладают влаголюбивые виды, зарегистрированные только во влажных или заболоченных лесах. Это Agonum gracile Gyll., обитающий в сосняке морошково-чернично-сфагновом. Виды Agonum micans Nic. и Platynus mannerheimii Dej., отмеченные в ельнике хвощово-папоротниковом. Виды Leistus terminatus Hellw., Patrobus assimilis Pk. приуроченные к влажным ельникам. Только на вершине холма в ельнике чернично-кислично-зеленомошном встречен вид Harpalus quadripunctatus Dej.
Отмечена неравномерность распределения вида Calathus micropterus Duft. вдоль линии профиля в течение сезона. Установлена повышенная активность имаго вида на вершинах и склонах холмов весной и осенью, а летом - в понижениях рельефа.
Обсуждение результатов
Сведения о богатстве фауны жужелиц коренных еловых и сосновых лесов Вологодской области подтверждают мнение о том, что по числу их видов подзоны тайги слабо различаются [30]. Так, для хвойных лесов в южной, средней и северной тайге авторами указывается от 22 до 32 видов. В то время как число видов в населении жужелиц биоценозов от южной к северной подзонам сокращается более существенно. Так по числу видов жужелиц отдельные хвойные биоценозы средней тайги (7 - 18 видов) занимают промежуточное положение между лесными сообществами южнотаежной (22 - 32 вида) и северотаежной (12 - 13 видов) подзон.
В составе фауны жужелиц лесных среднетаежных биоценозов Вологодской области преобладают европейско-сибирские и трансевразийские виды, что соответствует расположению изученного района в пределах равнинной местности центральной части европейской тайги. Кроме того, эта черта характерна для фауны жужелиц таежной зоны в целом и неоднократно отмечалось различными авторами [2, 10 др.].
Фауна жужелиц исследованного массива типична для влажных хвойных таежных биоценозов. Доминирующие и большинство редких в исследованном районе видов широко распространены в таежной зоне и являются многочисленными. Внимания заслуживает находка более южного вида Pterostichus quadrifoveolatus Letzn., которая свидетельствует о переходном характере фауны Шалго-Бодуновского леса. Это определено тем, что изученный массив расположен вблизи границы биомов средней и южной тайги.
Анализ комплексов жужелиц лесных биотопов Шалго-Бодуновского заказника показал, что их видовое богатство связано с особенностями почвенно-растительного покрова. Это зависимость неоднократно отмечалась многими отечественными и зарубежными карабидологами. Общепризнанно, что влияние растительного покрова на комплексы жужелиц опосредуется формированием микроклимата [27, 32, 33, 34 и др.]. В то же время растительность служит индикатором эдафических особенностей биотопа, которые также оказывают влияние на жужелиц. Среди них, считается, что наибольшее влияние на распределение почвенных беспозвоночных оказывает кислотность, поскольку она является интегральным показателем особенностей почв. Так, установлено, что параметры кислотности коррелируют с содержанием органического вещества в почве и ее влагоудерживающей способностью [22]. Условия увлажнения и кислотность почв изменяются в ряду изученных местообитаний (рис. 1). Важно, что по мере снижения кислотности происходит увеличение трофности почв (содержания элементов питания). Показатели трофности играют большую роль для формирования растительного покрова.
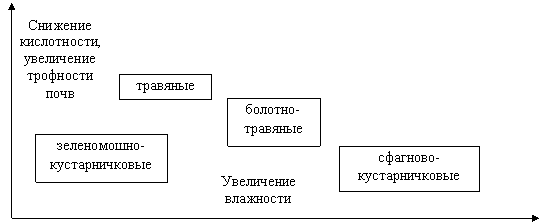
Рис. 1. Экологическая структура лесов Шалго-Бодуновского ландшафтного заказника (по: Ценофонд лесов... [28])
Зеленомошно-кустарничковые: сосняк-ельник чернично-зеленомошный, ельник чернично-кислично-зеленомошный. Травяные: ельник папоротниково-таволговый, ельник хвощово-папоротниковый, ельник хвощово-папоротниково-кисличный. Болотно-травяные: ельник травяно-болотный. Сфагново-кустарничковые: сосняк морошково-чернично-сфагновый
В биотопах с застойным режимом увлажнения, высокой кислотностью и низкой трофностью почв, а также покровом из сфагновых мхов и кустарничков (сосняк морошково-чернично-сфагновый) формируются комплексы жужелиц с самым низким видовым богатством. Также малое число видов отмечено для биотопов с нормальным увлажнением, кислыми, бедными элементами питания почвами и покровом из зеленых мхов и кустарничков (сосняк-ельник чернично-зеленомошный). Повышение видового богатства комплексов жужелиц отмечается в биоценозах, расположенных на влажных или временно избыточно увлажненных, слабокислых почвах с высокой трофностью и с хорошо развитым травянистым или травяно-кустарничковым покровом (травяные и болотно-травяные ельники). Таким образом, формированию высокого видового богатства комплексов жужелиц способствуют нормальное или временно избыточное увлажнение, низкая кислотность и высокая трофность почв, а также хорошо развитый травянистый покров. Значимость травяно-кустарничкового яруса для формирования комплексов жужелиц уже отмечалась для лесных сообществ северной тайги [30].
О численности жужелиц в биоценозах судят по показателям их уловистости, считается, что между данными параметрами существует прямая связь [11, 24, 30 и др.]. Поэтому, увеличение уловистости жужелиц в биотопах может трактоваться как повышение их численности, а снижение - как сокращение количества жуков в сообществах. В то же время, известно, что показатели уловистости жужелиц проявляют зависимость от их активности [14]. Таким образом, в изученных биотопах сокращение уловистости жужелиц в комплексах с высоким уровнем видового богатства может быть объяснено снижением их численности в сообществах в результате ужесточения межвидовых конкурентных взаимоотношений. Это правило широко известно в экологии [20]. Или же сокращение уловистости жужелиц может быть определено снижением активности жуков. Важно, что уловистость жужелиц сокращается именно в тех биотопах (ельник болотно-травяный и ельник хвощово-папоротниковый), где складываются условия, благоприятные для их существования (низкая кислотность, высокая влажность, хорошо развитый травянистый покров). Причин способствующих понижению активности жуков в такой ситуации может быть несколько. Экспериментально установлено, что в своих перемещениях жужелицы руководствуются поиском оптимальных абиотических условий (освещение, температура, влажность) [17]. Следовательно, если условия в биотопах благоприятны, активность, а значит и уловистость, жужелиц понижаются. В то же время для некоторых напочвенных животных (в частности для мокриц) лабораторно установлено снижение активности в условиях агрегированного расположения питательных ресурсов [22]. Значит, снижение активности жужелиц в биотопах возможно при формировании для них богатой кормовой базы. В пользу этого предположения в нашем исследовании свидетельствует установленный факт, что условия, благоприятные для жужелиц, оптимальны для напочвенных организмов в целом, в том числе и для их жертв [33]. Таким образом, в лесных биотопах с большим числом видов жужелиц для них имеется богатая кормовая база, что устраняет необходимость поиска пищи, а, следовательно, активность и уловистость.
В целом, следует отметить, что имеющихся данных пока недостаточно для корректного вывода о связи численности, активности и уловистости жужелиц в среднетаежных лесных сообществах. Однако, эти показатели, по-видимому, проявляют зависимость от микроклиматических особенностей биотопов, обилия пищи и напряженности межвидовых отношений.
Анализ видового состава комплексов жужелиц показывает, что большую роль в его формировании играет мозаичность расположения местообитаний различного типа. Об этом свидетельствует сходство фаун жужелиц в биотопах, располагающихся вдоль линии профиля на вершинах холмов и в межхолменных понижениях и граничащих между собой. Для них отмечено около 50% общих видов. Это определяется высокой динамической активностью жужелиц. Как уже было отмечено, жуки активно перемещаются в поисках благоприятных условий и пищи, в том числе и по линии профиля. Поэтому приуроченность видов к определенным местообитаниям выражена слабо. Для некоторых из них (например, Calathus micropterus Duft.) перемещения вдоль линии профиля имеют сезонный характер. Возможно, что в условиях мозаичного рельефа на вершинах холмов протекает зимовка имаго жужелиц, а в межхолменных понижениях они питаются. Однако выявлено, что некоторые виды жужелиц обитают только в пониженных участках рельефа или значительно реже на вершинах и склонах холмов.
Выводы
Таким образом, фауна жужелиц коренных среднетаежных лесов Вологодской области включает 27 видов из 13 родов. Это укладывается в представления о составе локальных лесных таежных фаун, но меньше общего числа видов, зарегистрированных в лесных биоценозах средней тайги. По составу изученная фауна типична для влажных хвойных таежных лесов, но отличается наличием некоторых южных видов, что характеризует ее как переходную между южной и северной тайгой. К числу доминирующих относятся виды, широко распространенные в лесных сообществах умеренных широт.
Пространственное распределение жужелиц по среднетаежным лесным биотопам определяется их биологическими особенностями и мозаичностью условий территорий. Так высокая подвижность жуков, а также сезонные миграции видов, определяют их перемещение и стирают грани между комплексами жужелиц различных биотопов. В то время как видовое богатство жужелиц среднетаежных лесных биоценозов проявляет зависимость от кислотности и влажности почв, а также развития травянистого покрова в занимаемых биотопах.
Население жужелиц лесных биотопов средней тайги характеризуется переходными чертами от их комплексов в биоценозах южной тайги к сообществам северной подзоны. Это выражается в промежуточных по значениям показателях видового богатства и уловистости жужелиц. Причем отмечено значительное сходство комплексов жужелиц лесных среднетаежных и северотаежных сообществ по показателям уловистости.
СПИСОК ЛИТЕРАТУРЫ:
- 1. Алалыкина Н., Целищева Л. // Вестн. Ин-та биологии Коми НЦ УрО РАН. 2005. № 2 (88). С. 16.
- 2. Арнольди К.В. // Животный мир СССР. М.: Изд-во АН СССР, 1953.
Т. 4. С. 434. - 3. Белова Ю.Н. // Актуальные проблемы биологии и экологии: Матер. докл. XII Молодежн. конф. Ин-та биологии Коми НЦ УрО РАН. Сыктывкар, 2006. С. 29.
- 4. Белова Ю.Н. // Международный контактный форум по сохранению местообитаний в Баренцевом регионе. Четвертое совещание. Матер. докл. Сыктывкар, 2005. С. 254.
- 5. Болотова Н.Л., Шабунов А.А., Коновалов А.Ф. // «Проблемы особо охраняемых природных территорий Европейского Севера (К 10-летию Национального парка «Югыд Ва»)»: Матер. научно-практич. конф. Сыктывкар, 2004. С. 48.
- 6. Воронин А.Г. // Энтом. обозр. 2000. № 2. С. 328.
- 7. Гиляров М.С. Зоологический метод диагностики почв. М.: Наука, 1965. 280 с.
- 8. Грюнталь С.Ю. // Проблемы почвенной зоологии. Минск: «Наука и техника», 1978. С. 304
- 9. Грюнталь С.Ю. // Вестн. зоологии. 1981. № 5. С. 20.
- 10. Грюнталь С.Ю. // Фауна и экология почвенных беспозвоночных Московской области. М.: Наука, 1983. С. 85.
- 11. Грюнталь С.Ю. // Бюл. МОИП, отд. биол. 1996. Т. 101. Вып. 1. С. 50.
- 12. Грюнталь С.Ю., Бутовский Р.О. // Энтом. обозр. 1997. Т. LXXVI. Вып. 3.
С. 547. - 13. Грюнталь С.Ю. // Изв. РАН. Сер. Биологич. 1998. № 2. С. 292.
- 14. Душенков В.М. // Экология жизненных форм почвенных и наземных членистоногих. М., 1986. С. 25.
- 15. Еремин П.К. // Кологривский лес (Экологические исследования). М., 1986. С. 110.
- 16. Ермаков А.И. // Экология. 2004. № 6. С. 45.
- 17. Кауфман Б.З., Бобровских Т.К. // Энтом. обозр. 1991. Т. LXX. №1. С. 23
- 18. Красная книга Карелии. Петрозаводск: Карелия, 1995. 286 С.
- 19. Крылова Л.П. // Экологические аспекты сохранения видового разнообразия на европейском северо-востоке России. Сыктывкар, 1996. С. 90.
- 20. Одум Ю. Экология. М.: Мир. 1986. Т. 1. 328 с.; Т. 2. 364 с.
- 21. Перель Т.С. // Pedobiologia. 1965. № 5. С. 102.
- 22. Покаржевский А.Д., Гонгальский К.Б., Зайцев А.С., Савин Ф.А. Пространственная экология почвенных животных / Отв. ред. Добровольский Г.В. М.: Товарищество научных изданий КМК, 2007. 174 с.
- 23. Разнообразие ландшафтов национального парка «Русский Север» / Ред.: Н.К. Максутова. Вологда, 2007. 92 С.
- 24. Рыбникова И.А. // Тр. Дарвин. гос. природного биосферного заповедника. Череповец, 2006. Вып. XVI. С. 166.
- 25. Рыбникова И.А., Власов Д.В., Видягина Е., Белова Ю.Н. // Биологические ресурсы Белого моря и внутренних водоемов Европейского Севера: Сб. матер. IV (XXVII) Междунар. конф. Ч. 2. Вологда, 2005. С. 104.
- 26. Ужакина О.А. // Каталог жуков комплексного заказника «Белоярский». Сыктывкар, 2002. С. 21.
- 27. Феоктистов В.Ф., Душенков В.М. // Зоол. журн. 1982. Т. LXI. Вып. 2. С. 227.
- 28. Ценофонд лесов Европейской России // http://mfd.cepl.rssi.ru/flora/type_forest2.htm.
- 29. Шарова И.Х. // Фауна и экология животных. М.: МГПИ им. В.И. Ленина, 1971. С. 61. 30. Шарова И.Х., Филиппов Б.Ю. Экология жужелиц лесов в дельте Северной Двины: Монография. Архангельск: Поморский университет, 2004. 116 с.
- 31. Яковлев А.И. // Тр. Ярослав. естественно-исторического общ-ва. Ярославль, 1902. Т. 1. С. 88.
- 32. Brose U. // Oecologia. 2003. Vol. 135. P. 407.
- 33. Butterfield J., Luff M.L., Baines M., Eyre M.D. // Forest Ecology and Management. 1995. Vol. 79. P. 63.
- 34. Ings T.C., Hartley S.E. // Forest Ecology and Management. 1999. Vol. 119.
P. 123. - 35. Lindroth C.H. // Fauna entomologica Scandinavica. 1985. Vol. 15, part 1. 205 p.
Библиографическая ссылка
Белова Ю.Н. КОМПЛЕКСЫ ЖУЖЕЛИЦ КОРЕННЫХ СРЕДНЕТАЕЖНЫХ ЛЕСОВ (НА ПРИМЕРЕ ВОЛОГОДСКОЙ ОБЛАСТИ) // Современные проблемы науки и образования. – 2009. – № 3. ;URL: https://science-education.ru/ru/article/view?id=1135 (дата обращения: 17.04.2024).