



Семейство Rosaceae - сложная в систематическом отношении группа. Традиционно в семействе выделялось 4 подсемейства: Rosoideae, Spiraeoideae, Maloideae и Amygdаloideae (Prunoideae). Однако молекулярно-генетические исследования изменили представление о системе этого семейства. В системе Rosaceae, опубликованной D. Potter в 2007 году [10], объем подсемейства Amygdаloideae увеличился. Сюда были отнесены подсемейства Spiraeoideae и Maloideae. Таким образом, положение многих родов в подсемействах Rosaceae в системах, основанных на молекулярно-генетических исследованиях, не всегда совпадает с таковым в системах, основанных на анатомо-морфологических признаках. Следует отметить, что спорная ситуация в семействе Rosaceae существовала и раньше. Особенно различались взгляды на положение отдельных видов и родов. Например, В.С. Симагин [3-5], занимающийся изучением морфологии косточковых плодовых растений, разделял виды черешни и вишни Cerasus avium, C.vulgaris, C.fruticosa и Microcerasus tomentosa по отдельным родам. Цвелев [7] и Камелин [1] выделили M.tomentosa в самостоятельный род. В системе Поттера [10], основанной на молекулярно-генетических исследованиях, эти виды отнесены к одному роду Prunus.
Для уточнения таксономического статуса родов и видов необходимо привлечение дополнительных, ранее не использующихся в систематике растений признаков. Так как мужские генеративные структуры наиболее вариабельны по сравнению с женской сферой, использование их эмбриологических характеристик в качестве диагностических признаков может внести вклад в систематику и филогению родов семейства Rosaceae.
Материал и методы исследования
Объектами исследования были представители наиболее крупных и широко распространенных во флоре Евразии родов двух подсемейств (по: Potter et. al., 2007): наиболее архаичное - Spiraeoideae (трибы Sorbarieae, Pyreae, Spiraeeae, Amygdаleae, Neillieae) и Rosoideae (трибы Sanguisorbeae, Colurieae, Potentilleae). Наиболее детально мужские генеративные структуры были изучены у представителей триб Amygdаleae и Spiraeeae. Характеристика пыльников представителей остальных триб дорабатывается.
Строение стенки микроспорангия у исследуемых видов изучали на постоянных микропрепаратах, изготовленных по общепринятой методике [2]. Материал окрашивали гематоксилином по Гейденгайну.
Результаты исследования и их обсуждение
Изучение стенки микроспорангия проводилось на стадии зрелой пыльцы. Пыльник у изученных видов 4-гнездный, с лартрорзным вскрыванием. Стенка микроспорангия развивается по типу однодольных в классификации Davis [9], или центростремительному типу в классификации Терехина и др. [6]: у всех видов клетки париетального слоя, являющиеся производными клеток археспория, делятся периклинально. При этом наружный слой дифференцируется в эндотеций, а клетки внутреннего слоя делятся с образованием среднего слоя и тапетума. Таким образом, в стенке микроспорангия выделяются 4 слоя: эпидерма, эндотеций, средний слой и тапетум [8].
Несмотря на сходный план строения стенки микроспорангия, форма и размеры клеток каждого слоя у разных видов различаются даже в пределах одного рода.
Наименее вариабельными из всех слоев стенки микроспорангия являются клетки эпидермы. У всех изученных видов клетки эпидермы слабо вакуолизированы, вытянутой формы. У представителей разных видов лишь немного различается форма клеток. Так, у представителей близких видов, относящихся, согласно классификации Potter et. al. [10] к одному роду Prunus в трибе Amygdаleae, - вишни степной и вишни обыкновенной (Cerasus vulgaris, C. fruticosa) клетки эпидермы более широкие, с ровными очертаниями, тогда как у представителей того же рода - вишни войлочной (Microcerasus tomentosa) они слегка угловатые, у черешни (C.avium) клетки эпидермы почти квадратные, миндаля низкого (Amygdalus nana) - в основном неправильной формы (рис. 2, 1-6). У видов трибы Spiraeeae клетки эпидермы также мало различаются (рис. 1, 1-3).
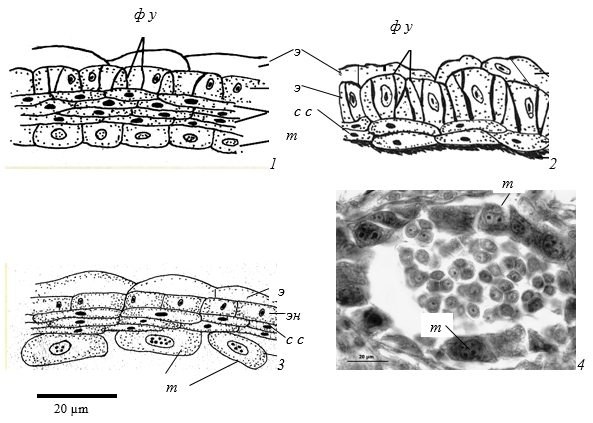
Рис. 1. Стенка зрелого микроспорангия у представителей подсемейства Spiraeoideae
1 - Physocarpus amurensis, 2 - Sorbaria sorbifolia, 3 - Spiraea chamaedryfolia; 4 - Physocarpus amurensis
э – эпидерма, эн – эндотеций, с с - средний слой, т - тапетум.
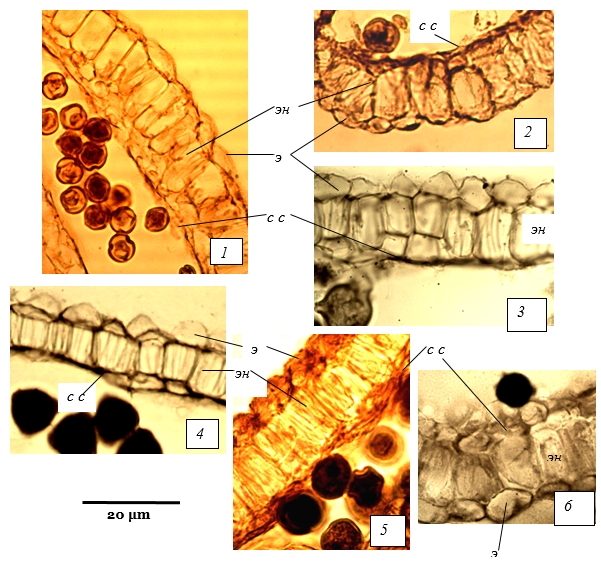
Рис. 2. Стенка зрелого микроспорангия у представителей подсемейства Rosoideae.
1 - Cerasus vulgaris, 2 - C. fruticosa, 3 - Amygdalus nana, 4 - Microcerasus tomentosa, 5, 6 - Cerasus avium; э - эпидерма; эн - эндотеций; с с - средний слой
У видов более всего различаются клетки эндотеция. На начальных стадиях развития пыльника в клетках эндотеция образуются мелкие вакуоли, ядро занимает центральное положение. Постепенно вакуоли объединяются, и ядро смещается к внутренней тангентальной стенке. В клеточных стенках эндотеция у всех видов имеются фиброзные утолщения. Они позволяют пыльнику вскрываться продольными щелями в области стомиума. Толщина фиброзных поясков неодинаковая. Например, у Sorbaria sorbifolia и Physocarpus amurensis фиброзные утолщения значительные – в виде большой капли (рис. 1, 1, 2). У многих других видов Rosaceae фиброзные утолщения небольшие или вообще не видны.
Сильно варьируют форма и размеры клеток эндотеция. Например, у C.avium в зрелом пыльнике клетки эндотеция вытянуты в высоту, широкие, с ровными очертаниями (рис. 2, 5, 6). В то же время, у A.nana клетки напоминают многоугольники, неправильной формы (рис. 2, 3). У части видов, характеризующихся крупными клетками эндотеция, некоторые его клетки дополнительно делятся. В этих местах эндотеций становится дву- или трехрядным. Поделившиеся клетки впоследствии не увеличиваются.
Средний слой. В начале формирования стенки микроспорангия в среднем слое один ряд клеток. Затем за счет периклинальных делений число рядов увеличивается. Деления клеток среднего слоя неупорядоченные, поэтому в стенке одного микроспорангия можно наблюдать в разных местах неодинаковое число рядов клеток – 1-5 с дистальной стороны микроспорангия и 2-7 рядов – с латеральных сторон. Ко времени образования зрелой пыльцы клетки среднего слоя сильно сжимаются и сминаются. Из-за этого на срезах пыльника иногда сложно подсчитать число их рядов. У представителей разных видов количество средних слоев разное. Например, на стадии тетрад микроспор у Physocarpus мы наблюдали 3-4 слоя, Sorbaria 2 слоя, Spiraea – 2-3 слоя (рис. 1, 1-3), M.tomentosa - 2, C.avium – 3 слоя (рис. 2, 4-6).
Тапетум, который идет на питание развивающихся микроспор, однорядный у всех изученных видов, клеточный, без реорганизации. У Spiraea и Physocarpus клетки тапетума расходуются постепенно, видны даже на стадии зрелой пыльцы (рис. 1, 3, 4). В то же время, у Sorbaria, Cerasus, Amygdalus и др. в зрелом пыльнике тапетум полностью расходуется (рис. 2, 1-6).
Таким образом, у представителей двух изученных триб семейства Rosaceae форма и размеры клеток слоев стенки микроспорангия сильно варьируют. Эти признаки могут быть использованы в систематике растений Rosaceae. Сравнительная эмбриологическая характеристика пыльников разных видов в комплексе с другими анатомо-морфологическими признаками растений может решить и некоторые спорные вопросы филогении. Представители подсемейств Spiraeoideae и Rosoideae в разных системах относят к наиболее архаичным группам, из которых спирейные считают наиболее древними. Это подтверждают и данные нашего эмбриологического исследования. Так, большое число слоев среднего слоя свидетельствует о примитивности вида. Наибольшее число слоев среднего слоя имеется у видов Spiraea. Наличие выраженных фиброзных утолщений в стенках клеток эндотеция, скорее всего, следует отнести к прогрессивному признаку. Фиброзные утолщения облегчают механизм вскрывания пыльника, доступ опылителей. Следует отметить, что выраженные фиброзные утолщения мы наблюдали у S.sorbifolia и Physocarpus amurensis, отнесенных к наиболее архаичной группе. Такое несоответствие объясняется явлением гетеробатмии, при которой специализация структур растения в процессе эволюции происходит с неодинаковой скоростью. В итоге у примитивных по внешним морфологическим признакам групп могут встречаться более прогрессивные цитоэмбриологические признаки. В этом случае для решения вопросов филогении требуется привлечение большего числа признаков.
Заключение
Результаты проведенного исследования показывают, что для уточнения таксономического статуса и филогении представителей видов Rosaceae следует привлекать эмбриологические характеристики мужских генеративных структур. Таксоноспецифичными признаками пыльника являются число рядов клеток среднего слоя, форма и размеры клеток эндотеция, наличие или отсутствие фиброзных утолщений в клетках эндотеция, присутствие клеток тапетума в сформированной стенке микроспорангия.
Библиографическая ссылка
Яндовка Л.Ф. ТАКСОНОСПЕЦИФИЧНЫЕ ОСОБЕННОСТИ СТРОЕНИЯ ПЫЛЬНИКА У ПРЕДСТАВИТЕЛЕЙ СЕМЕЙСТВА ROSACEAE // Современные проблемы науки и образования. – 2016. – № 6. ;URL: https://science-education.ru/ru/article/view?id=25824 (дата обращения: 24.04.2024).