



Одним из наиболее приоритетных направлений в клеточной биологии и молекулярной медицине является изучение механизмов эпигенетической регуляции функций органов и тканей на клеточном уровне. Сохранение гомеостазиса и биологического единства организма на тканевом, субклеточном и клеточном уровне возможно за счет действия регуляторных белков и пептидов, реализующих многоступенчатое равновесное состояние между различными физиологическими процессами - пролиферацией, дифференцировкой, запрограммированной гибелью клеток. Многочисленные данные, описанные в этой области знаний, способствуют пониманию фундаментальных механизмов молекулярно-клеточного гомеостазиса, обмена и воспроизведения генетической информации и позволяют экстраполировать эти заключения для поиска инновационных подходов к диагностике, профилактике и терапии заболеваний, а также повышения продолжительности жизни.
Перспективным и научно подтвержденным экспериментальным методом увеличения продолжительности и качества жизни может служить использование коротких геропротекторных пептидов, содержащих 2-4 аминокислотных остатка. Пептиды Ala-Glu-Asp-Gly, Glu-Asp-Gly, Ala-Glu-Asp-Lys, Lys-Glu активируют экспрессию мРНК генов роста, развития и дифференцировки культуры клеток растения табака (Nicotiana tabacum) [1]. У мутантной линии дрозофилы agnts3 пептид Glu-Asp-Arg улучшает двигательную активность и процессы запоминания информации путем эпигенетического воздействия на экспрессию мРНК генов limk1, rok, park. Мутации в этих генах являются ключевыми факторами развития болезни Паркинсона. Введение пептидов Ala-Glu-Asp-Gly и Lys-Glu в культуру клеток эмбрионов сетчатки цыплят активирует дифференцировку различных типов нейронов и пигментного эпителия сетчатки глаза [2]. Наибольшее повышение продолжительности жизни животных на 42,3% было отмечено при введении пептида Ala-Glu-Asp-Gly. При применении ДНК-микрочиповой технологии был изучен эффект пептидов Lys-Glu, Glu-Trp, Ala-Glu-Asp-Gly, Ala-Glu-Asp-Pro на экспрессию 15247 генов сердца и головного мозга мышей [3]. Пептиды Ala-Glu-Asp-Gly, Lys-Glu, Ala-Glu-Asp, Lys-Glu-Asp индуцируют нейрональную дифференцировку стволовых клеток тканей зуба человека (hPDLSCs) и являются перспективными для изучения в качестве модуляторов нейрогенеза [4]. Пептиды специфически регулируют экспрессию определенной группы генов и синтез соответствующих белков. У старых обезьян (Macaca mulatta) после инъекционного введения пептидов эпифиза выявлена нормализация уровня мелатонина и кортизола в крови [3]. Применение пептидов в различных экспериментальных моделях приводило к повышению/понижению экспрессии мРНК генов и нормализации синтеза белков, кодируемых этими генами, что коррелировало с повышением функций различных органов и тканей и повышением продолжительности жизни.
Известно, что 20 кодируемых аминокислот, являющихся структурными элементами белков и пептидов, регулируют активность различных органов и тканей [5]. При уменьшении концентрации экстрацеллюлярного глютамина у клеток увеличивается восприимчивость к Fas-опосредованной запрограммированной клеточной гибели [6]. Этот эффект также может служить одной из причин дисфункции скелетных мышц [7]. В литературе описаны данные о регуляции пролиферации и запрограммированной гибели клеток под влиянием аминокислоты аргинина. Введение фермента, снижающего уровень аргинина за счет энзиматической деградации (аргиназы), в культуру нормальных клеток способствовало аресту клеточного цикла в фазах G0 и G1 с его последующей нормализацией. Следует отметить, что в иммортализованной клеточной культуре при введении аргинина способность клеток к пролиферации не восстанавливалась [8]. Лейцин, аминокислота с разветвленными боковыми радикалами, в диапазоне концентраций от 10–5 до 10–3 M повышал синтеза ДНК и пролиферации гепатоцитов. Более того, лейцин повышал активность фермента S6-киназы 1 (S6K1) и эукариотического фактора инициации (eIF4E) [9]. Ранее нами было показано, что кодируемые аминокислоты активируют либо ингибируют пролиферацию и запрограммированную гибель клеток в эксплантатах тканей животных разного возраста [10].
Целью работы стало изучение совместного действия коротких пептидов Ala-Glu-Asp-Pro, Ala-Glu-Asp-Arg, Lys-Glu-Asp-Ala, Glu-Asp-Pro и Lys-Glu-Asp и концевых аминокислот, в структуре данных пептидов, на развитие эксплантатов тканей коры головного мозга, сердца, печени, селезенки и сосудов животных разного возраста.
Материал и методы исследования. В исследовании было использовано более 700 эксплантатов тканей селезенки, печени, сердца, коры головного мозга, 3-месячных (молодых) и 20-месячных (старых) крыс-самцов линии Вистар из коллекции лабораторных млекопитающих разной таксономической принадлежности вивария Института физиологии им. И.П. Павлова РАН. Выделенные кусочки тканей диспергировали на более мелкие фрагменты размером 1 мм3. Эти фрагменты культивировали в чашках Петри в питательной среде, в состав которой входило 35% среды Игла, 35% раствора Хенкса, 25% фетальной бычьей сыворотки. В культуральную среду добавляли глюкозу (0,6%), инсулин (0,5 ед/мл) и антибиотик гентамицин в количестве 100 ед/мл. Все эксплантаты разделяли на 3 группы: 1 – контрольная, с 3 мл питательной среды, без добавления пептидов и аминокислот, 2 – в культуры вводили по 3 мл питательной среды с исследуемой концентрацией пептидов и 3 – в культуры вводили по 3 мл питательной среды с исследуемой концентрацией пептидов и аминокислот. Эксплантаты выращивали в СО2-инкубаторе при температуре 37 °С при 5% СО2 в течение 3 суток. После этого образцы культур изучали под фазово-контрастным микроскопом. Пептиды кортаген (Ala-Glu-Asp-Pro), кардиоген (Ala-Glu-Asp-Arg), ливаген (Lys-Glu-Asp-Ala), кристаген (Glu-Asp-Pro), везуген (Lys-Glu-Asp) и добавляемые к ним аминокислоты (Sigma, США) добавляли в культуральную среду в конечных концентрациях 0,05 нг/мл. К пептиду Ala-Glu-Asp-Pro добавляли аминокислоту пролин (Pro), к пептиду Ala-Glu-Asp-Arg – аргинин (Arg), к пептиду Lys-Glu-Asp-Ala – аланин (Ala), к пептиду Glu-Asp-Pro - пролин (Pro), к пептиду Lys-Glu-Asp – аспарагин (Asp). Для изучения сочетанного действия пептидов и аминокислот были выбраны аминокислоты, содержащиеся на концах пептидов, т.к. в предварительных экспериментах именно они обладали наибольшей биологической активностью. На 3-и сутки выращивания вокруг эксплантатов появлялась зона роста, которая была образована пролиферирующими и выселяющими клетками фибробластоподобной морфологии. Для оценки зоны роста эксплантатов тканей использовали численный показатель – индекс площади зоны роста эксплантата (ИП). Рассчитывали среднее значение ИП по 15 эксплантатам. ИП рассчитывали, как отношение разницы площади всего эксплантата, совместно с периферической зоной роста, с вычетом центральной зоны, к площади центральной зоны эксплантата. ИП в группе «контроль» принимали равным 100%. ИП в исследуемой группе, с добавлением биологически активных веществ, выражали в процентах к контролю. Достоверность различий в группах оценивали по критерию Стъюдента в программе Statistica 7.0. Различия между группами считали статистически достоверными при уровне значимости р <0,05.
Результаты исследования и их обсуждение
Действие сочетанного действия пептидов и аминокислот, входящих в их состав, на органотипические культуры тканей молодых животных. Пептид Ala-Glu-Asp-Pro повышал рост эксплантатов коры головного мозга на 22% (n=15, p<0,05) в сравнении с этим показателем в группе «контроль» (n=14). Пептид Ala-Glu-Asp-Pro и аминокислота пролин повышали ИП эксплантатов коры головного мозга на 38% (n=15, p<0,05) по сравнению с этим показателем в группе «контроль». Эффект сочетанного действия пептида Ala-Glu-Asp-Pro и пролина на рост органотипической культуры ткани коры головного мозга молодых крыс был больше на 16%, чем действие одного пептида (рис. 1).
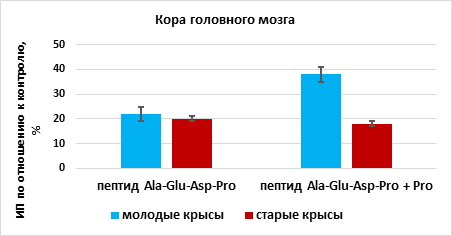
Рис. 1. Влияние пептида Ala-Glu-Asp-Pro и пролина (Pro) на рост органотипической культуры ткани коры головного мозга молодых и старых крыс
* - р< 0,05 по сравнению с соответствующим показателем в группе
«пептид Ala-Glu-Asp-Pro»
В присутствии пептида Ala-Glu-Asp-Arg происходила стимуляция роста органотипической культуры ткани сердца на 20% (n=13, p<0,05), по сравнению с этим показателем в группе «контроль» (n=15). При культивировании фрагментов сердца в присутствии пептида Ala-Glu-Asp-Arg и аргинина выявлено статистически значимое увеличение ИП эксплантатов на 41% (n=13, p <0,05) по сравнению с этим показателем в группе «контроль». Эффект сочетанного действия пептида Ala-Glu-Asp-Arg и аргинина на рост органотипической культуры ткани сердца молодых крыс был больше на 21%, чем действие одного пептида (рис. 2).
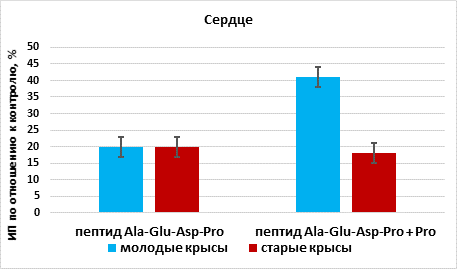
Рис. 2. Влияние пептида Ala-Glu-Asp-Arg и аргинина (Arg) на рост органотипической культуры ткани сердца молодых и старых крыс
* - р< 0,05 по сравнению с соответствующим показателем в группе
«пептид Ala-Glu-Asp-Arg»
Пептид Lys-Glu-Asp-Ala стимулировал рост эксплантатов печени на 20% (n=15, p <0,05) по сравнению с этим показателем в группе «контроль» (n=13). При культивировании фрагментов печени в присутствии пептида Lys-Glu-Asp-Ala и аланина происходило статистически значимое увеличение ИП эксплантатов на 35% (n=14, p <0,05) по сравнению с этим показателем в группе «контроль». Эффект сочетанного действия пептида Lys-Glu-Asp-Ala и аланина на рост органотипической культуры ткани печени молодых крыс был больше на 15%, чем действие одного пептида (рис. 3).
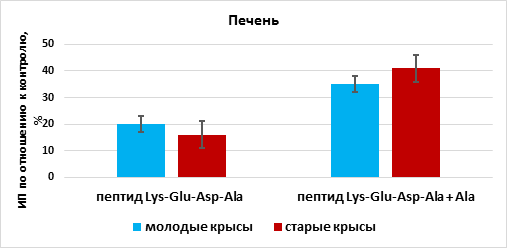
Рис. 3. Влияние пептида Lys-Glu-Asp-Ala и аланина (Ala) на рост органотипической культуры ткани печени молодых и старых крыс
* - р< 0,05 по сравнению с соответствующим показателем в группе
«пептид Lys-Glu-Asp-Ala»
В присутствии пептида Glu-Asp-Pro происходила стимуляция роста органотипической культуры селезенки на 22% (n=13, p<0,05) по сравнению с этим показателем в группе «контроль» (n=13). При культивировании фрагментов селезенки в присутствии пептида Glu-Asp-Pro и пролина установлено статистически значимое увеличение ИП эксплантатов на 31% (n=12, p <0,05) по сравнению с этим показателем в группе «контроль». Эффект сочетанного действия Glu-Asp-Pro и пролина на рост органотипической культуры ткани селезенки молодых крыс был больше на 9%, чем действие одного пептида (рис. 4).
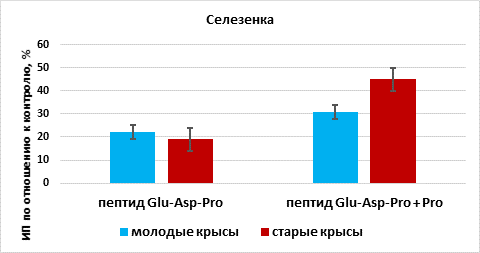
Рис. 4. Влияние пептида Glu-Asp-Pro и пролина (Pro) на рост органотипической культуры ткани селезенки молодых и старых крыс
* - р< 0,05 по сравнению с соответствующим показателем в группе «пептид Glu-Asp-Pro»
Пептид Lys-Glu-Asp стимулировал рост эксплантатов сосудов на 61% (n=15, p<0,05) по сравнению с этим показателем в группе «контроль» (n=13). При культивировании фрагментов сосудов в присутствии пептида Lys-Glu-Asp и аспарагина происходило статистически значимое увеличение ИП эксплантатов на 74% (n=12, p <0,05) по сравнению с этим показателем в группе «контроль». Эффекты сочетанного действия Lys-Glu-Asp и аспарагина на рост органотипической культуры ткани сосудов молодых крыс достоверно не отличались от действия одного пептида (рис. 5).
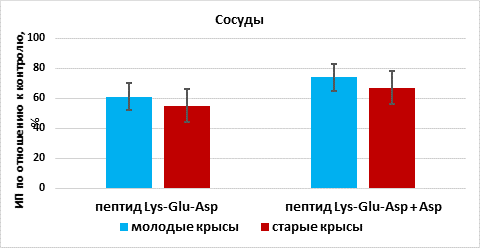
Рис. 5. Влияние пептида Lys-Glu-Asp и аспарагина (Asp) на рост органотипической культуры ткани сосудов молодых и старых крыс
Влияние сочетанного действия пептидов и аминокислот, входящих в их состав, на органотипические культуры тканей старых животных. Пептид Ala-Glu-Asp-Pro стимулировал рост эксплантатов коры головного мозга на 20% (n=16, p <0,05) по сравнению с этим показателем в группе «контроль» (n=15). При культивировании фрагментов коры головного мозга в присутствии пептида Ala-Glu-Asp-Pro и пролина наблюдалось статистически значимое увеличение ИП эксплантатов на 18% (n=16, p <0,05) по сравнению с этим показателем в группе «контроль». Таким образом, пептид Ala-Glu-Asp-Pro и сочетание этого пептида с пролином оказывали одинаковый стимулирующий эффект на рост эксплантатов коры головного мозга старых животных (рис. 1).
В присутствии пептида Ala-Glu-Asp-Arg происходила стимуляция роста органотипической культуры ткани сердца на 21% (n=17, p <0,05) по сравнению с этим показателем в группе «контроль» (n=14). При культивировании фрагментов сердца в присутствии пептида Ala-Glu-Asp-Arg и аргинина выявлено статистически значимое увеличение ИП эксплантатов на 19% (n=16, p <0,05) по сравнению с этим показателем в группе «контроль». Пептид Ala-Glu-Asp-Arg и сочетание этого пептида с аргинином оказывали одинаковый стимулирующий эффект на рост эксплантатов сердца старых крыс (рис. 2).
Пептид Lys-Glu-Asp-Ala стимулировал рост эксплантатов печени на 16% (n=18, p <0,05) по сравнению с этим показателем в группе «контроль» (n=15). При культивировании фрагментов печени в присутствии пептида Lys-Glu-Asp-Ala и аланина происходило статистически значимое увеличение ИП эксплантатов на 41% (n=18, p <0,05) по сравнению с этим показателем в группе «контроль». Увеличение ИП при сочетанном действии тетрапептида и аланина превосходило ИП при действии одного пептида Lys-Glu-Asp-Ala на 25% (рис. 3).
В присутствии пептида Glu-Asp-Pro происходила стимуляция роста ткани селезенки на 19% (n=15, p <0,05) по сравнению с этим показателем в группе «контроль» (n=15). При культивировании фрагментов селезенки в присутствии пептида Glu-Asp-Pro и пролина установлено статистически значимое увеличение ИП эксплантатов на 45% (n=15, p <0,05) по сравнению с этим показателем в группе «контроль». Увеличение ИП при сочетанном действии трипептида и пролина превосходило ИП при действии пептида Glu-Asp-Pro на 24% (рис. 4).
Пептид Lys-Glu-Asp стимулировал рост эксплантатов сосудов на 55% (n=16, p <0,05) по сравнению с этим показателем в группе «контроль» (n=16). При культивировании фрагментов сосудов в присутствии пептида Lys-Glu-Asp и аспарагина происходило статистически значимое увеличение ИП эксплантатов на 67% (n=15, p <0,05) по сравнению с этим показателем в группе «контроль». Таким образом, пептид Lys-Glu-Asp и сочетание этого пептида с аспарагином оказывали стимулирующий эффект на рост эксплантатов сосудов старых животных, статистически не различавшийся (рис. 5).
Заключение. В присутствии концевых аминокислот, содержащихся в пептидах Ala-Glu-Asp-Pro, Ala-Glu-Asp-Arg, Lys-Glu-Asp-Ala, Glu-Asp-Pro, усиливается стимулирующее действие пептидов на рост эксплантатов коры головного мозга, селезенки, печени и сердца молодых животных. Под действием пептидов ИП зоны роста указанных тканей повышался на 20-22%, тогда как при сочетанном действии пептидов и аминокислот этот показатель возрастал до 31-41%.
В присутствии концевых аминокислот, содержащихся в коротких пептидах Lys-Glu-Asp-Ala и Glu-Asp-Pro, повышается стимулирующее действие пептидов на рост органотипических культур тканей печени и селезенки старых крыс. Под действием пептидов ИП зоны роста указанных тканей повышался на 16-19%, тогда как при сочетанном действии пептидов и аминокислот этот показатель возрастал до 41-45%. Можно полагать, что при одновременном воздействии пептидов Lys-Glu-Asp-Ala и Glu-Asp-Pro и их концевой аминокислоты на ткани печени и селезенки старых крыс происходит суммирование эффектов этих двух биологически активных молекул. При этом в органотипических культурах коры головного мозга, сердца и сосудов старых животных не выявлено отличий по влиянию на площадь зоны роста тканей при добавлении к пептидам аминокислот. Это может быть связано с тем, что ткани коры головного мозга, сосудов и сердца менее восприимчивы к дополнительной стимуляции их пролиферативного потенциала под действием аминокислот.
В органотипических культурах ткани сосудов молодых и старых животных пептид Lys-Glu-Asp и его сочетание с аспарагином в одинаковой степени стимулировали рост эксплантатов – на 61-74%. Вероятно, отсутствие различий между действием пептида Lys-Glu-Asp и его сочетания с аминокислотой на рост органотипической культуры тканей сосудов связано с исходно высокой биологической активностью пептида.
Стимулирующее влияние сочетаний коротких пептидов с аминокислотами может быть обусловлено тем, что короткие пептиды могут проникать в цитоплазму и ядро клетки и связываться с ДНК. В то же время кодируемые аминокислоты обладают способностью регулировать специфические гены на транскрипционном и трансляционном уровнях, что способствует регуляции метаболизма клетки [11; 12].
Исследованные сочетания коротких пептидов Ala-Glu-Asp-Pro, Ala-Glu-Asp-Arg, Lys-Glu-Asp-Ala, Glu-Asp-Pro с концевыми аминокислотами открывают перспективы для создания новых лекарственных препаратов, стимулирующих функции тканей коры головного мозга, селезенки, печени и сердца при патологических процессах, в том числе ассоциированных с возрастом.
Библиографическая ссылка
Рыжак Г.А., Чалисова Н.И., Ивко О.М., Гутоп Е.О. СОЧЕТАННОЕ ДЕЙСТВИЕ КОРОТКИХ ПЕПТИДОВ И АМИНОКИСЛОТ НА РОСТ ОРГАНОТИПИЧЕСКИХ КУЛЬТУР ТКАНЕЙ МОЛОДЫХ И СТАРЫХ ЖИВОТНЫХ // Современные проблемы науки и образования. – 2019. – № 5. ;URL: https://science-education.ru/ru/article/view?id=29149 (дата обращения: 26.04.2024).