



В тундровой зоне Западно-Сибирской равнины отмечено 218 видов птиц, из них 138 гнездятся [8, 10]. Хорологический анализ гнездовой орнитофауны этой территории показал, что, согласно макроареалогической характеристике, наиболее значимую часть гнездящейся орнитофауны составляют относительно широко распространённые арктогейские голарктические и трансдолготно-палеарктические виды [10]. По зонально-ландшафтному распределению среди гнездящихся птиц преобладают виды гипоарктические, бореальные, температно-субтропические, температные, гемиарктические и эоарктические. При совмещении макроареалогической и зонально-ландшафтной характеристик наиболее многочисленны две группы трансдолготно-палеарктических видов (гипоарктические и температно-субтропические) и голарктические бореальные виды. Классификация локальных орнитофаун тундровой зоны Западно-Сибирской равнины показала, что наиболее значимая орнитофаунистическая граница проходит между южной и средней подзональными полосами субарктических тундр, и несколько менее значима граница между подзонами арктических и субарктических тундр [9].
Цель исследования – оценить территориальную неоднородность населения птиц тундровой зоны Западно-Сибирской равнины в период размножения, сравнить её с орнитофаунистической неоднородностью и выявить основные факторы, влияющие на формирование пространственно-типологической структуры и организации населения этой группы позвоночных животных с точки зрения методологии факторной зоогеографии [20, 21].
Материал и методы исследования
Учёты птиц проведены без ограничения ширины трансекта, с последующим пересчётом обилия на площадь по средним групповым дальностям обнаружения, рассчитанным интервальным методом [20]. Большинство учётов проведено на постоянных, но не строго фиксированных маршрутах. По каждому местообитанию данные усреднены за первую половину лета (16 июня – 31 июля). В целом для большей части населения птиц это время близко периоду гнездования [11]. Протяженность основных учётных маршрутов, включая литературные данные, составила 3516 км, дополнительных (когда учитывали отдельные виды) – 1003 км, всего 4519 км. Данные всех учётов птиц, в том числе на реках, озёрах и вдоль береговых линий морских заливов, приведены в числе особей на 1 км2. Опубликованные материалы по плотности населения птиц с полуострова Ямал использованы только в тех случаях, когда они были даны по отдельным местообитаниям, а не на территорию стационаров в целом [22, 23]. В тех случаях, когда данные по Ямалу представлены числом пар или гнёзд, эти показатели удвоены. С учётом литературных данных, материалы собраны в 1971–1974, 1978, 1981, 1983–1991, 2002, 2003, 2005–2008 гг., т.е. в течение 21 полевого сезона.
Тундровая зона Западно-Сибирской равнины делится на подзоны арктических и субарктических тундр, при этом вторая включает 3 подзональные полосы (северных моховых, низкокустарниковых и южных кустарниковых тундр) [12]. В настоящей статье подзональные полосы названы, соответственно, северная, средняя и южная. Координаты и сроки работ для большинства ключевых участков указаны ранее [4]. Кроме того, нами использованы данные по учётам птиц на других участках подзоны субарктических тундр (рис. 1).
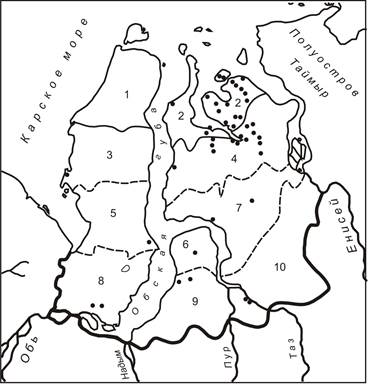
Рис. 1. Зонально-подзональное деление тундровой зоны Западно-Сибирской равнины и места сбора материала
Условные обозначения. Границы: ▬ - лесной и тундровой зон; — - подзон арктических и субарктических тундр; ------- - подзональных полос подзоны субарктических тундр.
Участки: Подзона арктических тундр: 1, 2. Подзона субарктических тундр: подзональная полоса северных моховых субарктических тундр: 3, 4; подзональная полоса низкокустарниковых субарктических тундр: 5, 6, 7; подзональная полоса южных кустарниковых субарктических тундр: 8, 9, 10. Участки 1, 3, 5 и 8 – полуостров Ямал; участки 2, 4, 7 и 10 – Гыданский полуостров; участки 6 и 9 – Тазовский полуостров.
Точками показаны места проведения учётов птиц.
Материалы обработаны с помощью пакета программ банка данных лаборатории зоологического мониторинга ИСиЭЖ СО РАН. Для расчётов использована матрица коэффициентов сходства вариантов населения птиц Жаккара [27] в модификации Р.Л. Наумова [17]. Формула коэффициента Жаккара – Наумова следующая: KJ-N = ΣC•100%/(А+B–ΣC), где ΣC – сумма меньших показателей обилия видов, общих для двух сравниваемых вариантов населения; A, B – плотность населения (суммарное обилие) всех видов в первом и втором вариантах населения. Для классификации вариантов населения птиц использована программа KLAFA, алгоритм которой представляет собой объединённый качественный вариант методов главных компонент и кластерного анализа [24]. Оценка связи факторов среды и неоднородности населения птиц сделана по выборке из 114 вариантов населения с помощью линейной качественной аппроксимации (аналога регрессионной модели) [19].
Фоновыми считали виды, обилие которых равно не менее 1 особи/км2. Доминантами считали виды, доля которых по тому или иному показателю составляет не менее 10 %. Первые 3 вида, наиболее значимые по какому-либо показателю, считали лидерами, вне зависимости от их доли. Для каждого класса указаны первые 3 лидера, а в случаях, когда 4-й вид является доминантом, указывали и его. Названия видов даны по списку птиц Северной Евразии в границах бывшего СССР [13] с некоторыми изменениями [6, 7, 26]. Виды чечёток обыкновенная Acanthis flammea и пепельная A. hornemanni во время учётов трудно различимы, поэтому их считали вместе под общим названием чечётка A. flammea. Всего в анализируемой выборке фигурирует 118 видов птиц.
Для каждого местообитания оценены характеристики среды: состав растительности, увлажнённость, обводнённость, рельеф, принадлежность подзоне или подзональной полосе, численность леммингов в год исследования, заболоченность и степень антропогенного влияния (экспертно, в баллах или номинально). Например, фактор "состав растительности” представлен градациями “пойменные леса”, “высокие кустарники”, “кустарники средней высоты”, “низкие кустарники”, “травяные местообитания”, “мохово-лишайниковые тундры” и “растительность населённых пунктов, берегов озёр и рек”. Каждый фактор отражен 0–1-й матрицей (в строках – варианты населения, в столбцах – градации фактора: единица в случаях присутствия фактора, ноль при его отсутствии). Мера значимости фактора или природного режима – доля учтённой при классификациях дисперсии коэффициентов сходства.
Для оценки фактора “численность леммингов” все варианты населения птиц разделили по годам на 4 группы: пик, спад и рост численности леммингов, а также данные, усреднённые за несколько лет по местообитаниям, вне зависимости от численности леммингов. Чаще всего цикл численности леммингов занимает 3 года. У обоих видов леммингов (сибирского Lemmus sibiricus и копытного Dicrostonyx torquatus) он протекает синхронно. С 1971 и 1985 гг. данные о численности леммингов взяты из литературы [2, 3]. За 1986–1991, 2000–2008 гг. мы использовали собственные учёты. За 1988–2008 гг. использованы данные по динамике численности грызунов с сайта “Arctic birds” [25]. Спады численности леммингов в тундровой зоне Западно-Сибирской равнины отмечены (указаны только годы сбора данных по населению птиц) в 1971, 1974, 1980, 1983, 1986, 1989, 2003, 2006 и 2008 гг. Рост численности леммингов зафиксирован в 1972, 1978, 1981, 1984, 1987 и 1990 гг. Пики численности пришлись на 1973, 1979, 1985, 1988, 1991, 1994, 2002, 2005 и 2007 (рост и пик) гг.
Результаты исследований и их обсуждение
Пространственно-типологическая структура орнитокомплексов выявлена на основе классификации населения с учётом максимальных значений сходства между выделенными классами и максимальным значением снятой дисперсии матрицы коэффициентов сходства. Наилучшей признана классификация из 8 таксонов, при которой снятая дисперсия составила 57 %, а коэффициент корреляции 0.7. Структурный граф представляет собой два параллельных вертикальных ряда классов, соединённых в верхней части рисунка, т.е. там, где отражены классы, включающие наиболее северные варианты населения птиц (рис. 2). Вертикальные ряды классов показывают отличия в населении птиц в зависимости от природной зональности территории – левый в местообитаниях суши, в том числе на болотах, правый на водоёмах и водотоках. Кратко полученная структура населения представлена нами ранее [5].
В верхней части рисунка эти два вертикальных ряда связывает 1-й класс, в который вошло население птиц водоёмов, водотоков и прибрежных (приморских) местообитаний суши подзоны арктических тундр. Население птиц прибрежных (приморских) местообитаний суши подзоны арктических тундр ближе орнитоценозам водоёмов и водотоков этой подзоны (1-й класс), а не к населению птиц суши (2-й класс). Это связано, прежде всего, с влиянием здесь приливов и штормов, что приводит к насыщению этих местообитаний водой и песком и делает их переходными между сушей и морем и, как следствие, к преобладанию по обилию водно-околоводных видов: кулика-воробья Calidris minuta, галстучника Charadrius hiaticula и морянки Clangula hyemalis.
Население птиц местообитаний суши средней полосы субарктических тундр распределилось между двумя классами. Орнитоценозы тундрового ландшафта этой полосы вошли в более северный, 2-й класс, куда входит также большинство проб населения местообитаний суши арктических и северных субарктических тундр (кроме населённых пунктов и их ближайших окрестностей). А население птиц пойм и болот средней полосы ближе населению птиц более южного класса, 3-го, т.е. орнитоценозам суши южных субарктических тундр (кроме пойменных лесов, населённых пунктов и их окрестностей). Эти две группы населения птиц (2-й и 3-й классы) разделились, в основном за счёт большего обилия в северной группе кулика-воробья и турухтана Philomachus pugnax, а в южной – берингийской трясогузки Motacilla tschutschensis, чечётки и белой куропатки Lagopus lagopus.
Сообщества птиц населённых пунктов и их ближайших окрестностей (5-й класс) оказались ближе всего населению птиц южной подзональной полосы субарктических тундр, а также пойм и болот средней полосы субарктических тундр (3-й класс). Эти два класса вариантов населения птиц связаны между собой благодаря сходному обилию берингийской трясогузки, краснозобого конька Anthus cervinus и желтоголовой трясогузки Motacilla citreola. Эти три вида, как и другие несинантропные виды, проникают на территорию населённых пунктов. Однако население птиц застроенных территорий выделяется в отдельный класс, в основном за счёт большего обилия белой трясогузки Motacilla alba, белохвостого песочника Calidris temminckii и пуночки Plectrophenax nivalis и меньшего - чечётки, белой куропатки, овсянки-крошки Ocyris pusillus и круглоносого плавунчика Phalaropus lobatus.
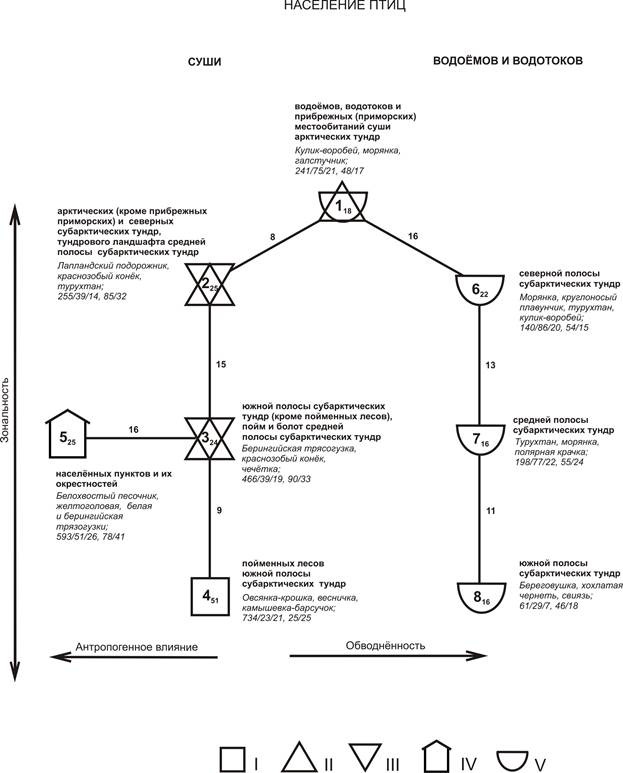
Рис. 2. Пространственно-типологическая структура населения птиц тундровой зоны Западно-Сибирской равнины в гнездовой период
Варианты населения разных групп местообитаний: I – лесов, II – тундрового ландшафта, III – пойм и болот, IV – застроенных территорий, V – водоёмов и водотоков. Направления изменений населения и увеличение проявления факторов показаны стрелками. Каждый значок соответствует группе объединившихся в один класс вариантов населения, при этом большая цифра означает номер группы, а индекс при ней – среднее сходство проб, вошедших в класс. Величины межклассового сходства населения указаны около каждой линии, соединяющей разные классы. Около каждого класса указаны группы вошедших в него вариантов населения птиц, первые 3-4 вида, преобладающих по обилию, а также 5 цифр (через косые черты), означающие следующие средние по классам значения параметров: суммарное обилие, особей/км2 / биомасса, кг / величина трансформируемой населением птиц энергии, ккал в сутки/км2 / общее число видов / число фоновых видов.
В пределах тундровой зоны Западно-Сибирской равнины наиболее сильно от остального населения птиц отличаются некоторые группы орнитоценозов южной подзональной полосы субарктических тундр: водоёмов и водотоков, и, особенно, пойменных лесов (соответственно, 8-й и 4-й классы). Это частично совпадает с орнитофаунистической неоднородностью исследуемого региона, т.к. наиболее сильные отличия в орнитофауне гнездящихся птиц отмечены между средней и южной подзональными полосами субарктических тундр [9].
Итак, при переходе от приморских прибрежных местообитаний суши арктических тундр и связанных с ними водоёмов и водотоков к югу в пределах местообитаний суши вплоть до пойменных лесов южной подзональной полосы субарктических тундр, вначале лидирующие по обилию кулик-воробей, морянка и галстучник уступают место лапландскому подорожнику Calcarius lapponicus, краснозобому коньку и турухтану, которые затем заменяются на берингийскую трясогузку и чечётку, которые, далее, уступают место овсянке-крошке, весничке Phylloscopus trochilus и камышевке-барсучку Acrocephalus schoenobaenus, а на застроенных территориях – белохвостому песочнику, желтоголовой, белой и берингийской трясогузкам. На водоёмах и водотоках, при переходе от подзоны арктических тундр к югу до южной полосы субарктических тундр, вначале лидирующие по численности кулик-воробей, морянка и галстучник заменяются на круглоносого плавунчика, турухтана и полярную крачку Sterna paradisaea, которые затем уступают место береговушке Riparia riparia, хохлатой чернети Aythya fuligula и свиязи Anas penelope.
В местообитаниях суши среди лидирующих по обилию видов преобладают представители отряда Воробьеобразные Passeriformes: лапландский подорожник, краснозобый конёк, берингийская, желтоголовая и белая трясогузки, чечётка, овсянка-крошка, весничка и камышевка-барсучок. На водоёмах и водотоках по обилию преобладают не воробьеобразные птицы: кулик-воробей, морянка, галстучник, круглоносый плавунчик, турухтан, полярная крачка, хохлатая чернеть и свиязь. Таким образом, основные тренды изменений населения птиц в тундровой зоне Западно-Сибирской равнины в гнездовой период связаны с обводнённостью и зональностью и, в меньшей степени, с антропогенным влиянием и облесённостью.
Видовое богатство. В среднем по населению птиц суши (классы 2–5-й) отмечено 70 видов птиц, а в населении птиц водоёмов и водотоков (классы 1-й, 6–8-й) только 51 вид. Это связано с двумя причинами. Во-первых, число водных видов птиц в нашей выборке меньше (38), чем число сухопутных видов (73), не считая видов, встречающихся в гнездовой период примерно с равной частотой в обоих типах местообитаний (7). Во-вторых, водоплавающих птиц нередко отмечают и на суше (к примеру, у гнёзд), в то время как большинство птиц суши не встречаются на воде. Они могут быть замечены только около воды, поэтому, как правило, не попадают в учёты птиц непосредственно на водоёмах и водотоках.
Суммарное обилие птиц. Максимальное по восьми классам суммарное обилие птиц отмечено в пойменных лесах южной полосы субарктических тундр (734 особи/км2), несмотря на то, что в этих вариантах населения фигурируют лишь гнездящиеся птицы. Однако, возможно, здесь наблюдается частичное завышение обилия при пересчёте опубликованных данных [23]. Дело в том, что это результат учётов птиц не по методике Ю.С. Равкина [20], как собрано большинство остальных анализируемых в данной статье материалов, а на площадках, выраженное в парах гнездящихся птиц. Возможно, при переводе этих данных в обилие особей/км2 (путём умножения числа пар на коэффициент 2), могло возникнуть завышение, к примеру, из-за того, что у ряда видов наблюдается полигиния или из-за других отличий двух указанных выше методик учёта птиц.
Минимальный показатель суммарного обилия птиц (61) отмечен в среднем по водоёмам и водотокам южной полосы субарктических тундр. В целом в местообитаниях суши с юга на север в пределах тундровой зоны в период гнездования прослеживается снижение суммарного обилия птиц. Это связано с ухудшением защитных условий в момент размножения из-за уменьшения высоты растительности, а также, видимо, с общим уменьшением продуктивности местообитаний суши из-за снижения теплообеспеченности. Однако на водоёмах и водотоках суммарное обилие к северу, напротив, как правило, увеличивается. Это объясняется тем, что антропогенное влияние с юга на север уменьшается. Оно сильнее сказывается на водоёмах и водотоках, чем в местообитаниях суши. Это связано с тем, что защитная функция относительно высокой растительности в южных тундрах выше, в то время как на водоёмах и водотоках птицы оказываются более уязвимыми к антропогенному воздействию. Некоторые формы антропогенного влияния (охота, браконьерство, рыбная ловля, судоходство) сильнее сказываются негативно именно на птицах водоёмов и водотоков южных участков тундровой зоны из-за большей плотности населения людей по сравнению с северными тундрами, а также из-за того, что хозяйственная деятельность людей проходит преимущественно в районах водных местообитаний.
В среднем по классам населения птиц водных местообитаний средняя масса одной особи птицы в 5 раз больше, чем в среднем по классам населения птиц суши (соответственно, 451 и 89 г.). При усилении антропогенного воздействия снижается численность, в первую очередь, крупных видов птиц. Так, на юге Ямала, в пределах южной полосы субарктических тундр в 1973 г., по сравнению с 1930-ми годами [15], из-за антропогенного влияния, уменьшилась, в первую очередь, численность ряда крупных видов птиц, в частности, тундрового лебедя Cygnus columbianus, белой совы Bubo scandiacus и сапсана Falco peregrinus. Сходное влияние антропогенных факторов на северо-востоке Азии привело к значительному сокращению численности ряда видов птиц, прежде всего, гусей: тундрового гуменника Anser serrirostris, белолобого гуся Anser albifrons и пискульки Anser erythropus [14].
Кроме того, в отличие от суши, общая продуктивность водных местообитаний, с юга на север, до известных пределов, увеличивается из-за того, что чем ниже температура воды, тем больше кислорода растворено в ней, в частности, в Обской губе [1]. Известно, что биологическая продуктивность поверхностных и приповерхностных вод напрямую зависит от объёма растворенного в воде кислорода. К примеру, летом объём первичной продукции в водах Обской губы с юга на север увеличивается, достигая максимума на границе речной и морской акваторий, которая находится в южной части подзоны арктических тундр, в районе посёлка Тамбей на западном берегу и мысов Хонарасаля и Штормовой – на восточном [16].
Суммарная биомасса птиц. Наибольший показатель суммарной биомассы птиц отмечен в среднем на водоёмах и водотоках северной подзональной полосы субарктических тундр (86 кг/км2). Наименьшие значения этого показателя характерны в среднем для двух классов южной полосы субарктических тундр: водоёмов и водотоков и пойменных лесов (23-29). Средняя суммарная биомасса населения птиц водоёмов и водотоков в гнездовой период почти вдвое больше (67), чем в среднем по местообитаниям суши (38).
Трансформируемая птицами энергия. Наибольший показатель величины трансформируемой птицами энергии характерен для населённых пунктов и их окрестностей (26 тыс. ккал. в сутки/кв. км), т.к. здесь отмечен один из самых высоких показателей суммарного обилия птиц. В большинстве остальных классов величина трансформируемой энергии ниже (14-22). Минимален этот показатель для населения птиц водоёмов и водотоков южной полосы субарктических тундр (7). В отличие от биомассы, в среднем по местообитаниям суши величина трансформируемой птицами энергии не меньше, а больше, чем таковая в среднем по водоёмам и водотокам, хотя и ненамного (соответственно, 20 и 18). Это связано с тем, что на суше существенно больше, чем на водоёмах доля особей отряда Passeriformes (уровень метаболизма которых выше, чем у птиц группы Non-Passeriformes).
Доля особей видов отряда Passeriformes. По восьми классам пространственно-типологической структуры населения птиц доля особей видов этого отряда снижается в двух направлениях: с юга на север и при переходе от урочищ суши к водоёмам и водотокам (рис. 3). Минимален этот показатель на водоёмах и водотоках от подзоны арктических тундр (10 %) до северной (2 %) и средней (6 %) полос подзоны субарктических тундр. Максимум отмечен в местообитаниях южной полосы субарктических тундр, пойм и болот средней полосы субарктических тундр (3-й класс) (72 %) и особенно в пойменных лесах южной полосы субарктических тундр (85 %). В среднем по местообитаниям суши доля особей отряда Passeriformes почти в 7 раз больше (66 %), чем в среднем по водоёмам и водотокам (10 %).
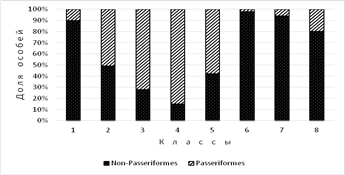
Рис. 3. Пространственные отличия доли особей отряда Passeriformes по классам пространственно-типологической структуры населения птиц тундровой зоны Западно-Сибирской равнины
Индекс интенсивности метаболизма населения птиц. Чтобы оценить, насколько интенсивно населением птиц трансформируется энергия (ккал) из расчёта на 1 г их суммарной биомассы, мы выяснили соотношение этих двух показателей (рис. 4). Наибольший индекс интенсивности метаболизма отмечен в пойменных лесах южной полосы субарктических тундр (0,97), так как в этом классе наибольшая доля особей видов отряда Passeriformes. Далее индекс падает в 5-м и 3-м классах (до 0,5), т.к. доля особей воробьеобразных птиц здесь заметно меньше. Во 2-м классе этот показатель уменьшается до 0,4, а наименьших значений он достигает в 1, 6, 7 и 8-м классах (0,2–0,3), т.к. доля особей отряда Passeriformes здесь наименьшая. В среднем по местообитаниям суши индекс интенсивности метаболизма населения птиц вдвое больше (0,6), чем в среднем по водоёмам и водотокам (0,3).
Средняя биомасса одной особи птиц. В пойменных лесах южной полосы субарктических тундр (4-й класс) отмечена наименьшая из восьми классов средняя биомасса одной особи – 31 г (рис. 5). Несколько больше этот показатель в местообитаниях южной полосы субарктических тундр (кроме пойменных лесов), поймах и болотах средней полосы субарктических тундр (3-й класс), а также в посёлках и их окрестностях (5-й класс, 83-85 г), и ещё больше в местообитаниях арктических (кроме прибрежных приморских) и северных субарктических тундр, а также в местообитаниях тундрового ландшафта средней полосы субарктических тундр (2-й класс, 156). И ещё вдвое выше этот параметр в среднем по водоёмам, водотокам и прибрежным приморским местообитаниям суши подзоны арктических тундр (1-й класс, 313). Максимальные значения характерны (по возрастанию) для водоёмов и водотоков средней, южной и северной полос субарктических тундр (соответственно, 386, 488 и 616 г). Таким образом, максимальный показатель средней биомассы одной особи по классам (616) в 20 раз больше минимального (31). В среднем по водоёмам и водотокам биомасса одной особи в 5 раз больше, чем среднем по местообитаниям суши (соответственно, 451 и 89 г).
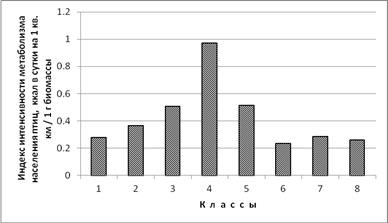
Рис. 4. Индекс интенсивности метаболизма населения птиц по классам пространственно-типологической структуры населения птиц тундровой зоны Западносибирской равнины в гнездовой период
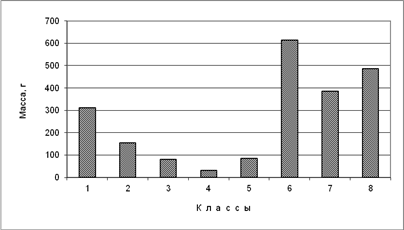
Рис. 5. Средняя масса одной особи птицы, по классам пространственно-типологической структуры населения птиц тундровой зоны Западносибирской равнины в гнездовой период
Средняя величина энергии, трансформируемая одной особью птицы. Минимальная величина энергии, трансформируемой одной особью птицы, отмечена в пойменных лесах южной полосы субарктических тундр (31 ккал в сутки / 1 кв. км) (рис. 6). Максимален этот показатель на водоёмах и водотоках всех подзональных полос субарктических тундр (110-143). В среднем по водоёмам и водотокам одной особью трансформируется почти втрое больше энергии (117 ккал), чем в среднем по суше (43), т.к. средняя масса одной птицы в среднем по водоёмам и водотокам в 5 раз больше, чем в среднем по местообитаниям суши.
Лидеры и доминанты по биомассе и трансформируемой птицами энергии чаще всего наиболее крупные по размерам виды птиц (табл. 1). В четырёх-пяти классах в число лидеров и доминантов по этим показателям входят морянка, в трёх – халей Larus heuglini и турухтан, в двух белолобый гусь, белая куропатка, только по биомассе ещё и морская чернеть Aythya marila. Остальные лидирующие по этим показателям виды отмечены лишь в какой-то одной группе местообитаний. По всем трём показателям (ещё и обилие) в трёх классах в число лидеров и доминантов входит морянка, в двух турухтан и в одном свиязь и хохлатая чернеть.
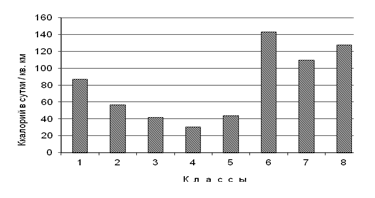
Рис. 6. Средняя величина энергии, трансформируемая одной особью птицы в сутки на 1 кв. км, по классам пространственно-типологической структуры населения птиц тундровой зоны Западно-Сибирской равнины в гнездовой период
Таблица 1
Лидеры и доминанты по обилию, биомассе и трансформируемой птицами энергии по классам пространственно-типологической структуры населения птиц тундровой зоны Западно-Сибирской равнины в гнездовой период
|
Класс |
Обилие, особей/км2 |
Биомасса, кг/км2 |
Трансформируемая энергия, ккал. в сут./км2 |
||||||
|
Всего |
Лидеры |
% |
Всего |
Лидеры |
% |
Всего |
Лидеры |
% |
|
|
1 |
241 |
Кулик-воробей Морянка Галстучник |
34 20 7 |
75 |
Морянка Халей Гага-гребенушка |
46 15 11 |
21 |
Морянка Халей Кулик-воробей |
42 11 11 |
|
2 |
255 |
Лапландский подорожник Краснозобый конёк Турухтан |
21 11 11 |
39 |
Белолобый гусь Белая куропатка Турухтан |
28 13 10 |
14 |
Турухтан Лапландский подорожник Белолобый гусь Белая куропатка |
14 12 11 10 |
|
3 |
466 |
Берингийская трясогузка Краснозобый конёк Чечётка |
17 12 9 |
39 |
Белая куропатка Шилохвость Морянка |
40 7 7 |
19 |
Белая куропатка Берингийская трясогузка Краснозобый конёк |
23 10 8 |
|
4 |
734 |
Овсянка-крошка Весничка Камышевка-барсучок |
19 13 8 |
23 |
Азиатский бекас Рябинник Бекас |
12 11 10 |
21 |
Овсянка-крошка Весничка Рябинник |
14 7 7 |
|
5 |
593 |
Белохвостый песочник Желтоголовая трясогузка Белая трясогузка Берингийская трясогузка |
13 12 10 10 |
51 |
Халей Турухтан Серая ворона |
32 8 6 |
26 |
Халей Турухтан Белохвостый песочник |
13 8 8 |
|
6 |
140 |
Морянка Круглоносый плавунчик Турухтан Кулик-воробей |
28 11 10 10 |
86 |
Морянка Белолобый гусь Морская чернеть |
32 19 11 |
20 |
Морянка Белолобый гусь Морская чернеть |
35 12 11 |
|
7 |
198 |
Турухтан Морянка Полярная крачка |
41 8 8 |
77 |
Халей Морянка Турухтан Морская чернеть |
17 15 15 11 |
22 |
Турухтан Морянка Халей |
27 14 12 |
|
8 |
61 |
Береговушка Хохлатая чернеть Свиязь |
13 11 10 |
29 |
Хохлатая чернеть Свиязь Морянка |
17 15 9 |
7 |
Хохлатая чернеть Свиязь Морянка |
16 14 9 |
Источники трансформируемой птицами энергии. Во всех классах пространственно-типологической структуры населения птиц энергия трансформируется птицами преимущественно за счёт беспозвоночных. Наиболее велика их доля в пойменных лесах южной полосы субарктических тундр (97 %). Минимально значение этого источника энергии на водоёмах и водотоках южной полосы субарктических тундр (60 %). В остальных классах этот показатель составляет 62–84 %. В среднем по водоёмам и водотокам доля беспозвоночных, в энергетическом эквиваленте, составляет 68 %, а в местообитаниях суши – 81 %. На суше больше, чем на водоёмах и водотоках доли, в энергетическом эквиваленте, беспозвоночных, а также семян и сочных плодов, но меньше – значение вегетативных частей растений и позвоночных.
Пространственная организация населения птиц
Наибольшую информацию о пространственной неоднородности населения птиц, выраженную долей учтённой дисперсии коэффициентов сходства (57 %), можно получить, классифицируя варианты населения, согласно полученной пространственно-типологической структуре (табл. 2). Чуть меньше дисперсии учитывается при делении территории и акватории на группы местообитаний, сходные с ландшафтами (53 %). В совокупности эти режимы дают 62 %. К этой оценке все факторы и природные режимы, при совместном их рассмотрении, в сумме добавляют лишь около 1 %, т.к. корреляция их влияния весьма велика.
Таблица 2
Доля дисперсии коэффициентов сходства населения птиц тундровой зоны Западно-Сибирской равнины (1974–2008 гг.), учтённой линейной качественной аппроксимацией при совокупной и индивидуальной оценке факторов и природных режимов, %
|
Фактор и режим |
Индивидуальная оценка |
Нарастающим итогом |
|
Состав растительности, в том числе значимость: травяного яруса лесов кустарников мохово-лишайниковых тундр плакора Увлажненность, в том числе значимость: близости водоёмов и водотоков многоводных пойм осоковых тундр болот заболоченных тундр сухих тундр Водность, в том числе значимость: рек и морских заливов рек микро-водоёмов и микро-водотоков озёр морских заливов Рельеф, в том числе значимость: пойм с водоёмами, без рек плакора, с понижениями плакора, без понижений понижений плакора долин с водоёмами Подзональность (подзоны и подзональные полосы) и провинциальность в том числе значимость: орнитофаунистической неоднородности (по [9]) южных субарктических тундр северных моховых субарктических тундр северных моховых и низкокустарниковых субарктических тундр арктических тундр, т.е. подзонального деления низкокустарниковых субарктических тундр Численность леммингов Заболоченность Антропогенное влияние |
48
27 19 14 7 38
24 9 4 3 2 0.8 27
12 8 5 5 2 21
14 8 7 1 0.5 14
11 9 3 2 1 0.8 8 7 2 |
48
56
57
59
60
61 61 61 |
|
Режимы По классификации населения Ландшафтное деление Всего (все учтённые факторы и режимы) |
57 53
|
62 62 63 |
При индивидуальной оценке факторов и природных режимов наиболее значим состав растительности. Этот фактор распадается на несколько градаций, из которых наиболее существенна роль травяного яруса. Меньше значимость лесов и кустарников, т.к. они, особенно леса, имеют в тундровой зоне меньшую площадь и, соответственно, меньшую долю в анализируемой выборке. Наиболее слабо в этой группе градаций проявляется значимость мохово-лишайниковых зональных тундр плакора, хотя их площадь в тундровой зоне велика. Это связано с заметным обеднением видового богатства обилия птиц в этом типе местообитаний.
Второе место среди комплексных факторов занимает увлажнённость. Из составляющих его наиболее значимо влияние водоёмов и водотоков, как среды, повышающей увлажнённость соприкасающихся с ними местообитаний суши. Слабее проявляется влияние сильно обводнённых пойм. Менее значимо влияют на неоднородность населения птиц отличия в увлажнённости осоковых тундр, болот и заболоченных тундр и ещё слабее – сухих тундр.
Увлажнённость значительно коррелирует с водностью, которая проявляется несколько слабее. Из градаций водности наиболее заметно совокупное влияние рек и морских заливов. При разделении этих режимов роль рек вчетверо выше значимости морских заливов. Отчасти это объясняется объёмами выборок по этим двум группам местообитаний. Немного меньшую, чем роль рек, и примерно одинаковую значимость имеют, с одной стороны, микро-водоёмы и микро-водотоки, с другой – озёра.
При оценке воздействия рельефа наиболее значимы отличия условий в поймах с водоёмами, без учёта рек. Роль плакора оценена дважды: с понижениями внутри него, т.е. с материковыми водоёмами и болотами, и без них. Во втором случае значимость плакора лишь немного меньше. Минимальна роль понижений плакора (материковых озёр и болот) и долин с водоёмами. Такая иерархия значимости этой группы природных режимов говорит о том, что население птиц понижений плакора и особенно долин рек не имеет существенных отличий от орнитоценозов сходных местообитаний пойм.
Корреляция неоднородность населения птиц с зональностью и подзональностью невелика. При разложении на составляющие наибольшее значение имеет деление орнитоценозов согласно орнитофаунистической классификации [9], т.е. в том случае, когда все население птиц поделено на три массива: орнитоценозы южной подзональной полосы субарктических тундр, подзоны арктических тундр и остальных территорий. К последним относятся две подзональные полосы подзоны субарктических тундр: северная и средняя.
Если оценивать каждую подзону и подзональную полосу отдельно, то выше всего значимость по населению птиц южной подзональной полосы субарктических тундр. Это совпадает с орнитофаунистической неоднородностью, согласно которой именно на границе между южной и средней подзональными полосами субарктических тундр проходит наиболее значимая орнитофаунистическая граница в тундровой зоне Западно-Сибирской равнины [9]. Однако, на следующем этапе орнитофаунистическая и населенческая неоднородности не совпадают. Так, согласно орнитофауне, следующая по значимости граница совпадает с подзональным геоботаническим делением тундровой зоны, т.е. проходит между подзонами арктических и субарктических тундр. А согласно неоднородности населения, после орнитоценозов южной подзональной полосы субарктических тундр, выделяется по значимости не подзона арктических тундр, а подзональная полоса северных моховых тундр. Это связано с тем, что для оценки орнитофаунистической неоднородности нами использованы не только собственные данные, но и опубликованные. Так, мы учитывали, что морской песочник Calidris maritimus гнездится только в подзоне арктических тундр, а в анализируемом массиве по населению птиц этого вида нет. Однако, в нашей выборке населению птиц есть 4 вида, отмеченных только в подзоне арктических тундр (белощёкая казарка Branta leucopsis, белоклювая гагара Gavia adamsii, плосконосый плавунчик Phalaropus fulicarius и моевка Rissa tridactyla), но их обилие незначительно. Поэтому, при оценке дисперсии коэффициентов сходства населения, вклад этих видов незначителен. Впрочем, в отличие от населения птиц южной полосы, значимость орнитоценозов подзоны арктических тундр и 2-х остальных подзональных полос субарктических тундр (северной и средней) в целом заметно меньше.
Влияние динамики численности леммингов равно 8 %, что, видимо, ниже реальной оценки, т.к. часть данных усреднена за годы с разной численностью леммингов. Меньше значимость заболоченности и, особенно, антропогенного влияния. Последнее связано с тем, что в посёлках тундровой зоны очень низко суммарное обилие птиц, обитающих только или преимущественно в населённых пунктах или их ближайших окрестностях (белая трясогузка, пуночка и полевой воробей Passer montanus). Эти виды связывают орнитоценозы населённых пунктов между собой и с таковыми их ближайших окрестностей. Однако, несинантропные виды, проникающие в населённые пункты (белохвостый песочник, желтоголовая и берингийская трясогузки) заметно больше сближают их орнитоценозы с населением птиц местообитаний, находящихся вдали от населённых пунктов.
Заключение
Ранее, при анализе пространственно-типологической структуры населения птиц всей Западносибирской равнины за 1971–1991 гг., в I половине лета в пределах тундровой зоны на уровне типов выделился только один тип населения, “тундровый”, куда вошли орнитоценозы всех тундровых местообитаний, кроме населения птиц водно-околоводных и застроенных местообитаний [18]. В этом типе населения первые три лидера по обилию – лапландский подорожник, берингийская трясогузка и краснозобый конёк. Население птиц водно-околоводных местообитаний тундровой зоны объединилось в один тип с населением сходных местообитаний лесотундровой зоны – “северный (тундрово-лесотундровый)”, где первые три лидера – турухтан, морянка и полярная крачка. Что касается населения птиц застроенной суши тундровой зоны, то оно ещё ближе орнитоценозам лесотундры и тоже объединилось с ним в один тип населения застроенной суши, названный “лесотундровым”. В нём лидируют по обилию белая трясогузка, береговушка и домовый воробей Passer domesticus.
Судя по пространственно-типологическим структурам населения птиц только тундровой зоны (настоящая статья) и всей Западно-Сибирской равнины [18], наиболее сильно население птиц тундровой зоны отличается от всего остального населения птиц Западно-Сибирской равнины в незастроенных местообитаниях суши, включая поймы и болота. Несколько меньше специфичность населения птиц водоёмов и водотоков тундровой зоны и ещё меньше она на застроенных территориях.
В проведении учётов птиц, кроме авторов, участвовали В.Г. Козин, П.М. Глазов, Н.Н. Емельченко, В.А. Юдкин, О.Д. Голубев, Н.Ю. Обухова, Д.С. Низовцев, Л.Г. Вартапетов, А.Е. Волков и Г.М. Тертицкий, которым авторы весьма благодарны. Мы благодарим также И.Н. Богомолову и Т.А. Кузнецову за помощь в статистической обработке данных и изготовлении рисунков.
Работа выполнена при поддержке РФФИ (гранты № 10-04-00639 и 10-04-00149-а).
Рецензенты:
Вартапетов Л.Г., д.б.н., зам. директора по науке, ФГБУН Институт систематики и экологии животных СО РАН, г. Новосибирск;
Литвинов Ю.Н., д.б.н., зам. директора по науке, ФГБУН Институт систематики и экологии животных СО РАН, г. Новосибирск.