



Вопросам эволюции нервной системы посвящено достаточное количество научных работ как российских, так и зарубежных авторов [1; 3]. Однако большинство авторов обращают внимание преимущественно на эволюционирование центральной нервной системы, а изменениям в периферической нервной системе уделяют незначительное внимание. Как известно, функциональной эволюции любой системы живого организма соответствует морфологическая эволюция. В доступной нам литературе достаточно подробно описаны морфологические изменения, наблюдаемые в центральной нервной системе при переходе позвоночных от водного к наземному образу жизни. Так, спинной мозг у амфибий развит значительно лучше, чем головной, состоящий из пяти отделов, расположенных в одной горизонтальной плоскости. В головном мозге наилучшего развития достигают промежуточный и продолговатый отделы. Мозжечок, в связи с однообразием движений и малой подвижностью, развит очень слабо [4; 6].
У ящериц, в сравнении с лягушками, эволюционные изменения в центральной нервной системе заключаются в следующем: передний мозг, состоящий из двух больших полушарий и имеющий абсолютно гладкую поверхность, сближается при помощи длинного обонятельного тракта с обонятельными луковицами, а не с промежуточными, как у амфибий. Передний мозг выполняет ведущую роль в организации поведения. Его размеры стали существенно больше и превосходят другие отделы головного мозга. У пресмыкающихся достаточно хорошо различим неопаллиум, который у земноводных выражен очень слабо. Мозжечок у ящериц более крупный и выпуклый в связи с разрастанием верхней поверхности. Его корпус развит сильнее, чем у лягушек. Теменной орган достигает своего максимального развития и функционирует как светочувствительный аппарат, реагирующий преимущественно на длинные световые волны. Что касается периферической нервной системы, то в области плечевого и тазового поясов образуются типичные нервные сплетения, обеспечивающие иннервацию скелетных мышц конечностей [5; 6]. Данные по периферической нервной системе, а именно по морфологическим перестройкам периферических нервов, скудные, однообразные и неполные [2; 7; 8]. Такое состояние проблемы и определило цель нашего исследования.
Цель исследования
Микроскопическое изучение морфологических особенностей периферической нервной системы в эволюционном аспекте на примере строения длинных ветвей плечевого сплетения в области средней трети плеча.
Материалы и методы
Исследование было проведено на достаточном количестве представителей класса земноводных (лягушка прудовая – Rana lessonae Camerano) и пресмыкающихся (ящерица прыткая – Lacerta agilis) (40 органокомплексов сосудисто-нервного пучка). Все исследуемые животные перед введением в эксперимент находились на двухнедельном карантине в экспериментально-биологической клинике (виварий) КГМУ, в течение которого были сформированы экспериментальные группы, рандомизированные по полу и весу. Обращение с животными, их содержание и умерщвление проводилось в соответствии с конвенцией «О защите позвоночных животных, используемых для экспериментов или в иных целях», принятой Советом Европы (Страсбург, 1986 г.), и директивой Совета 86/609/еес от 24.11.86 по согласованию законов, правил и административных распоряжений стран-участниц в отношении защиты животных, используемых в экспериментальных и других научных целях.
У исследуемых животных отсекали обе конечности плевого пояса, в области средней трети плеча производили забор сосудисто-нервных пучков нервов-сгибателей и нервов-разгибателей с окружающими мышцами. Полученный материал фиксировали в 10%-ном водном растворе нейтрального (кальциевого) формалина. Для общегистологического изучения полученный материал заливали в парафин по стандартной методике и микротомировали. Обзорное гистологическое исследование проводили на поперечных срезах сосудисто-нервных пучков, толщиной 10-12 мкм, окрашенных гематоксилином и эозином. Для изучения стромального компонента сосудисто-нервного пучка использовали окраску по Маллори и пикрофуксином по Ван-Гизону, железным гематоксилином по Гайденгайну. Для изучения проводникового компонента периферических нервов препараты окрашивали по методу Вейгерта-Паля (окраска миелиновых оболочек), толуидиновым синим по Нисслю. Для анализа полученных гистологических препаратов проводилась их микроскопия и описательная морфология. Морфометрия проводилась на цифровых микрофотографиях, полученных с помощью оптической системы микроскопа Leica-CME и окулярной фотонасадки DCM-510. Данные, полученные в условных единицах (пикселях), были переведены в абсолютные единица (мкм) с помощью специально выведенных коэффициентов для различных увеличений микроскопа.
На цифровых микрофотографиях поперечных срезов сосудисто-нервных пучков определяли количество первичных нервных пучков, образующих нерв-сгибатель или нерв-разгибатель; по кариологическим признакам определяли соотношение разных типов клеток, в окружающей нервные стволы соединительной ткани; измеряли площадь поперечного сечения сосудисто-нервного пучка окружающей соединительной ткани, нервных стволов; максимальные и минимальные диаметры нервных пучков; толщину периневрия, эндоневрия, миелиновой оболочки; подсчитывали количество миелиновых и безмиелиновых нервных волокон в нервных пучках и их соотношение, выраженное в относительных единицах (%). Полученные при исследовании морфометрические данные обрабатывали вариационно-статистическими методами. Для всех ранее названных параметров определяли минимальное и максимальное значения, среднюю арифметическую, ошибку средней арифметической. Достоверность различий определяли с помощью непараметрического критерия Вилкоксона-Манна-Уитни. При этом различия считали достоверными при 95%-ном пороге вероятности (Р≤0,05). Все вычисления выполнялись с помощью аналитического пакета приложения Excel Office 2010, лицензией на право использования которой обладает КГМУ.
Результаты исследования и их обсуждение
Сосудисто-нервные пучки периферических нервов плечевого сплетения в области средней трети плеча на поперечных срезах имели треугольную (нервы-сгибатели) или веретеновидную форму (нервы-разгибатели) как у лягушек, так у ящериц. Снаружи все сосудисто-нервные пучки были заключены в собственные тонкие фасциальные влагалища и образованы нервными стволами и крупными кровеносными сосудами артериального и венозного русла, между которыми располагалась соединительная ткань, обеспечивающая тесную взаимосвязь между компонентами сосудисто-нервного пучка и объединяющая их в анатомически и функционально единое целое (рис. 1).
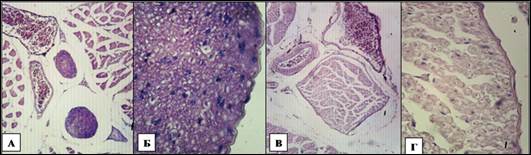
Рис. 1. Микрофотография сосудисто-нервных пучков периферических нервов плечевого сплетения в области средней трети плеча у земноводных и пресмыкающихся. У ящерицы - А, у лягушки - В. Ув. 200. Периневрий, эндоневрий и нервные волокна у ящерицы - Б, у лягушки - Г. Ув. 400.Окр. Г+Э.
Площадь поперечного сечения сосудисто-нервных пучков было достоверно (р≤0,5) больше в нервах-сгибателях у лягушки, а у ящерицы в нервах-разгибателях. При этом площадь поперечного сечения соединительной ткани, окружающей нервные пучки и крупные сосуды, имела ту же тенденцию, и ее средние значения в нервах-разгибателях у ящерицы были в 2,5 (на левой конечности) или 3,5 раза (на правой конечности) больше, чем в нервах-сгибателях (табл. 1).
Таблица 1
Средние значения некоторых показателей сосудисто-нервного пучка в области средней трети плеча у земноводных и пресмыкающихся
|
Показатели
Животные |
Площадь |
Площадь |
Площадь |
|
|
лягушка |
Левый сгиб. |
19,7±4,51* |
11,6±0,71* |
3,9±1,42* |
|
Левый разгиб. |
16,1±2,50* |
10,8±0,52* |
3,7±0,16* |
|
|
Правый сгиб. |
19,9±7,42* |
12,2±1,23* |
3,9±0,81* |
|
|
Правый разгиб. |
15,7±3,95* |
11,2±0,51* |
3,3±0,64* |
|
|
ящерица |
Левый сгиб. |
1,9±0,11* |
1,4±0,11* |
0,2±0,05* |
|
Левый разгиб. |
4,9±0,19* |
3,7±0,34* |
0,4±0,06* |
|
|
Правый сгиб. |
1,8±0,15* |
1,3±0,12* |
0,2±0,07* |
|
|
Правый разгиб. |
7,2±0,36* |
4,4±0,31* |
0,8±0,08* |
|
* р≤0,05 при сравнении конкретного параметра между экспериментальными животными.
У лягушки периферические нервы в области средней трети плеча состояли из одного-двух пучков в нервах-разгибателях и двух-четырех в нервах-сгибателях. У ящерицы количество нервных пучков было значительно больше и составило в нервах-разгибателях 3-4 пучка, в нервах-сгибателях 4-6 пучков, в зависимости от конечности. Нервные пучки у лягушки были овальной формы, у ящерицы преимущественно сферической или шарообразной формы. Максимальные значения площади поперечного значения у лягушки составили 3,99±1,42 мм2, а у ящерицы – 0,80±0,08 мм2 (табл. 1). Соотношение площади нервного пучка к площади, занимаемой соединительной тканью, было достоверно (р≤0,05) больше в 1,5-2 раза у лягушки, чем у ящерицы. При этом при вычислении соотношения площади поперечного сечения соединительной ткани к площади поперечного сечения сосудисто-нервного пучка достоверных отличий между животными выявлено не было. Этот показатель составил 0,64 у лягушки и 0,69 у ящерицы.
Интересно отметить, что нервные пучки как нерва-сгибателя, так и нерва-разгибателя у ящерицы располагались в эпиневрии не разрозненно, как у лягушки, а группами – по 2-3 пучка, объединенных общим тонким фасциальным футляром. В каждой такой группе нервных пучков присутствовал кровенаполненный магистральный сосуд.
Все нервные пучки окружены хорошо выраженным периневрием, в прослойках которого у ящериц встречаются мелкие кровеносные сосуды. Толщина периневрия и у лягушки, и у ящерицы существенно больше в нервах-сгибателях и составила соответственно 23,8±4,6 и 8,76±0,6 мкм слева и 22,6±3,9 и 5,2±0,6 мкм справа. Соотношение толщины периневрия к площади поперечного сечения нервного пучка было достоверно бо́льшим у ящерицы, что свидетельствует о более значимом развитии этой оболочки у пресмыкающихся. Нервные пучки образованы миелиновыми и безмиелиновыми нервными волокнами, между которыми расположены прослойки рыхлой волокнистой соединительной ткани эндоневрия. В эндоневрии ящериц встречается большее количество мелких кровеносных сосудов в стандартном поле зрения, чем у лягушки, при этом нервные волокна расположены более компактно и плотно.
Одним из важнейших направлений эволюции нервных элементов является миелинизация нервных волокон у позвоночных, которая существенно повышает скорость передачи нервного импульса. В нашем случае при подсчете количества миелиновых и безмиелиновых волокон в нервных пучках и определении их соотношения было выявлено, что количество миелиновых волокон у лягушки варьировало от 53 до 66% без достоверных отличий между нервами-сгибателями и нервами-разгибателями; у ящериц также преобладали миелиновые волокна, но их количество было значительно бо́льшим и варьировало от 68 до 90%, при этом достоверно (р≤0,05) бо́льшие значения наблюдались в нервных пучках нервов-сгибателей. Интересно также отметить, что в обеих экспериментальных группах диаметр миелиновых волокон был чуть больше диаметра безмиелиновых волокон, при этом наибольшие значения диаметра миелинового волокна наблюдались в нервных пучках нерва-сгибателя, преимущественно на левой конечности. Толщина миелиновой оболочки в мякотных волокнах имела похожую тенденцию, но при этом достоверно (р≤0,05) бо́льшие значения были выявлены в нервных волокнах, образующих нервы-сгибатели у пресмыкающихся (рис. 2).
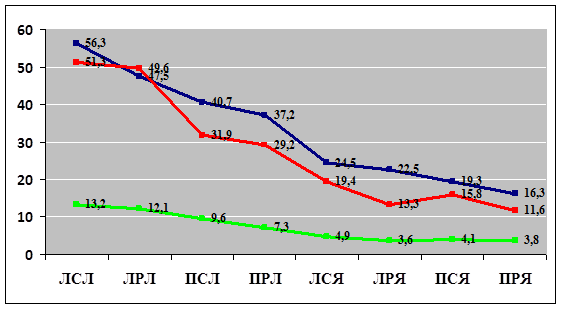
Рис. 2. Изменения диаметра нервных волокон (миелиновые волокна – синего цвета, безмиелиновые – красного) и толщины миелиновой оболочки (зеленого цвета) периферических нервов плечевого сплетения, выраженные в мкм, в области средней трети плеча правой и левой конечности у земноводных и пресмыкающихся.
Заключение. Таким образом, проведя комплексное гистологическое изучение периферических нервов в области средней трети плеча у животных, находящихся на разных ступенях эволюционной лестницы, мы выявили, что эволюция периферической нервной системы заключается как в качественном, так в количественном преобразовании структурных элементов нервного пучка. Основными морфологическими признаками эволюционирования периферической нервной системы у пресмыкающихся, в сравнении с земноводными, были следующие: изменение площади эпиневрия, окружающего нервные пучки; изменение диаметра и количества крупных кровеносных сосудов; увеличение количества нервных пучков в нервах-сгибателях и нервах-разгибателях; увеличение толщины периневрия и плотности расположения нервных волокон в эндоневрии на фоне увеличения количества кровеносных судов в нем. Увеличение относительного количества миелиновых нервных волокон и степени выраженности миелиновой оболочки. Наиболее ярко выраженные эволюционные преобразования периферических нервов происходят в нервах-разгибателях, возможно, в связи с выходом на сушу и появлением большего разнообразия движений. Полученные нами данные носят фундаментальный характер и дополняют знания о закономерностях эволюционных процессов периферической нервной системы.
Рецензенты:
Иванов А.В., д.м.н., профессор, заведующий кафедрой гистологии, цитологии, эмбриологии, ГБОУ ВПО «КГМУ Минздрава России», г. Курск;
Харченко В.В., д.м.н., профессор, заведующий кафедрой анатомии человека, ГБОУ ВПО «КГМУ Минздрава России», г. Курск.