



Как известно, одной из важных проблем в лесном хозяйстве Урала являются инфекционные болезни древесного яруса. Вследствие многократного порослевого возобновления и интенсивного антропогенного воздействия пораженность гнилевыми болезнями березняков в лесостепи Зауралья очень высока (60-85%) [4,5].
В целом, несмотря на интенсивные исследования в этом направлении, биохимические аспекты резистентности к гнилевым болезням и взаимодействия в системе: «патоген-растение-хозяин» остаются недостаточно выясненными, так как одни авторы отмечают накопление фенольных соединений в зоне некроза, вызванного инокуляцией патогенов, другие – их снижение [6-8,12].
Наиболее распространенным подходом в исследованиях является изучение биохимических процессов при искусственной инокуляции патогенов в зоне инокуляции. Ряд авторов отмечает, что некоторые фенольные соединения обладают выраженной фунгицидной активностью [10,11]. Имеются также единичные сообщения о том, что микозы у древесных растений сопровождаются активизацией синтеза фенольных соединений, детерминирующих параметры конститутивной резистентности [9]. Таким образом, в целом имеющиеся данные противоречивы. Исходя из этого, нами проведены исследования влияния поражения березы повислой стволовыми гнилями на состав и содержание фенольных соединений в листьях.
Материалы и методы
Для хроматографического анализа проводился сбор листьев березы повислой в Каменск-Уральском р-не Свердловской обл. Пробные площади располагались в березняках вблизи пос. «Покровское». Взятие каждой пробы осуществлялось от нескольких деревьев. Затем пробы смешивались для получения средней пробы. Непосредственно перед отбором проб определяли наличие или отсутствие пораженности дерева стволовыми и корневыми гнилями и стадию развития болезни с помощью взятия кернов из ствола и 3 корневых лап и их анализа. Как показали исследования, в припоселковых березовых лесах средний уровень антропогенной трансформации высок (4 стадия) [2]. Основной источник антропогенного воздействия: рекреационная и пастбищная дигрессия. Контрольные пробы отбирали от здоровых деревьев, не пораженных гнилевыми и другими болезнями, опытные пробы брали с деревьев, идентифицированных нами как пораженные стволовыми гнилями. Отбор проб проводился на одних и тех же пробных площадях, от деревьев одинакового диаметра, растущих в одинаковых лесорастительных условиях. Сразу после сбора листья березы высушивали при 600С, затем размалывали. После этого навеску с 2 г размолотых листьев смешивали с 20 мл 95% этанола. Экстракцию фенольных соединений из листьев березы проводили в обратном холодильнике на водяной бане в течение 30 мин. при кипении раствора. Затем суспензию центрифугировали при 10000 g в течение 10 мин. Супернатант фильтровали через фильтр с диаметром пор 0,2 мкм. Хроматографию проводили на жидкостном хроматографе Shimadzu LC-20 со спектрофотометрическим УФ детектором. Детектирование элюента осуществляли одновременно на двух полосах поглощения: 254 и 360 нм на колонке PerfectSil Target ODS-35мкм с обращенной фазой.
Градиентное элюирование проводилось в диапазоне 10–50 % со скоростью 1 мл в мин. при температуре 400С. Элюент А – ацетонитрил – 0,05 М фосфатный буферный раствор (рН=3,0); элюент В – ацетонитрил-вода (9:1). Продолжительность хроматографического анализа – 45 мин при 4000С. Из них от 0 до 30 мин. проводилось градиентное элюирование в диапазоне 10-50%, затем в течение 15 мин. при концентрации 50%. Для идентификации фенольных соединений использовали вещества-свидетели фирмы: Fluka, Sigma, Aldrich: аскорбиновая к-та, арбутин, галловая к-та, гиперозид, гидрохинон, кверцетин, изокверцетин, кофейная к-та, мирицетин, рутин, салицин, кафтаровая к-та, рутин; феруловая к-та, 3-КФК, 4-КФК, 5-КФК, кемпферол.
Результаты
Градиентный хроматографический анализ (ВЭЖХ) образцов из листьев березы повислой, взятых как с деревьев, пораженных стволовой гнилью, так и здоровых, позволил выявить 57 фенольных соединений (рис. 1).
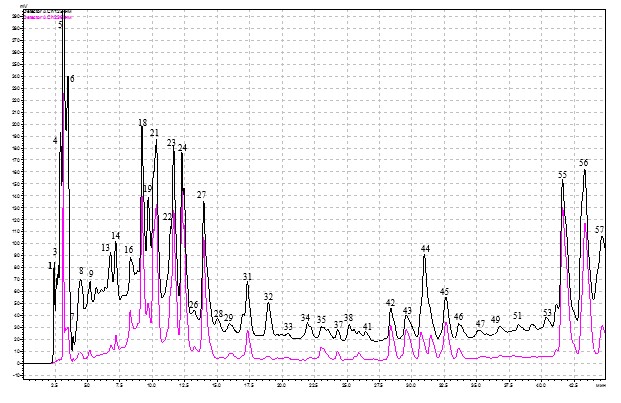
Рис. 1. ВЭЖХ фенольных соединений листьев березы повислой, пораженных стволовой гнилью. По оси абсцисс- tr (время удерживания), ординат-оптическая плотность
Примечания: 1 – аскорбиновая кислота; 4 – арбутин, 5 – галловая кислота; 7 – кафтаровая кислота; 8 – салицин, 13 – 4-кофеоилхинная кислота; 18 – рутин, 21 – 5-кофеоилхинная кислота; 23 – гиперозид; 24 – изокверцетин; 31 – мирицетин; 40 – кверцетин; 43 – кемпферол
Из общего количества фракций нами идентифицировано 13 фенольных соединений. Список идентифицированных фенольных соединений в листьях березы представлен в табл. 1. По химическому составу это были флавоноиды, фенолгликозиды, фенолкарбоновые и гидроксикоричные кислоты.
Для анализа основных, доминирующих тенденций влияния стволовой гнили на содержание фенольных соединений в листьях березы повислой нами проведен как тотальный сравнительный количественный попарный анализ основных выявленных при хроматографии пиков, так и сравнительный анализ идентифицированных нами фенольных соединений (табл. 1).
Таблица 1
Влияние стволовой гнили на состав и содержание фенольных соединений в листьях березы повислой
|
№ п/п |
Наименование |
Контроль Абсорбция (254 нм), mv |
Стволовая гниль Абсорбция (254 нм), mv |
Изменение содержания соединений (в % к контр.) |
Соотношение Y360 нм/ Y 254 нм |
ВУ |
|
1 |
Аскорбиновая к-та |
110,09 |
101,0 |
-8,18 |
0,0008 |
2,64 |
|
2 |
Арбутин |
285,0 |
193,0 |
-32,28 |
0,075 |
3,06 |
|
3 |
Галловая к-та |
90,5 |
237,0 |
+161,88 |
0,021 |
3,6 |
|
4 |
Салицин |
58,3 |
48,0 |
-17,24 |
0,0001 |
4,29 |
|
5 |
Кафтаровая к-та |
24,5 |
12,1 |
-50,61 |
0,095 |
4,003 |
|
6 |
4-кофеоилхинная к-та |
29,01 |
46,0 |
+36,93 |
0,30 |
6,7 |
|
7 |
Рутин |
43,0 |
113,0 |
+61,95 |
0,98 |
9,46 |
|
8 |
5-кофеоилхинная к-та |
90,06 |
143,0 |
+58,0 |
0,65 |
10,0 |
|
9 |
Гиперозид |
293,37 |
177,5 |
-39,50 |
0,884 |
10,93 |
|
10 |
Изокверцетин |
39,75 |
140,0 |
+252,2 |
1,00 |
11,74 |
|
11 |
Мирицетин |
22,5 |
44,2 |
+96,44 |
1,04 |
17,05 |
|
12 |
Кверцетин |
6,0 |
9,1 |
+51,66 |
0,97 |
25,4 |
|
13 |
Кемпферол |
21,8 |
20,5 |
--- |
1,22 |
28,5 |
Тотальный попарный сравнительный анализ образцов листьев пораженных и контрольных древостоев (пиков с идентичным ВУ и спектральным соотношением (S360 /S254)) убедительно продемонстрировал, что эти образцы значительно отличались между собой. Так, у 35,9% фракций их содержание заметно увеличивалось, у такого же количества уменьшалось, у 28,2% не изменялось. Для более глубокого анализа биохимических механизмов, происходящих в клетках листьев березы в насаждениях, пораженных стволовой гнилью, было целесообразно проанализировать тенденции реакций идентифицированных нами на хроматограммах фенольных соединений, так как функции ряда из них хорошо известны.
Как показали результаты исследований, у 50% идентифицированных нами фенольных соединений их содержание заметно увеличивалось. Это галловая к-та (161,88%), 4-кофеоилхинная к-та (36,93%), рутин (61,95%), 5-кофеоилхинная к-та (58,0%), изокверцетин (252,2%), мирицетин (96,44%) и кверцетин (51,66%). Обращает внимание значительный уровень возрастания активности галловой к-ты, изокверцетина и мирицетина. Одновременно снижается уровень активности аскорбиновой к-ты, арбутина, салицина, кафтаровой к-ты, гиперозида. Сравнение полученных нами результатов с имеющимися в литературе не дает положительных результатов, так как нами изучались биохимические изменения на значительной стадии пораженности, когда площадь пораженной древесины превышала 30% по диаметру, тогда как другие авторы изучали чаще биохимический состав после непосредственной инокуляции патогена. Так, синтез таких фенолгликозидов, как арбутин и салицин не активируется, но, в целом общий уровень активности идентифицированных соединений значительно возрастает.
Учитывая то, что большинство идентифицированных нами фенольных соединений являются активными антиоксидантами, можно заключить, что биохимические процессы борьбы с оксидативным стрессом у березы, пораженной стволовой гнилью, заметно активизируются. Вместе с тем, известно, что некоторые патогенные грибы сами выделяют антиоксиданты для разрушения Н2О2 и супероксидного радикала, которые выполняют защитную функцию, способствующую выживанию патогена (Абрамова и др., 2008). Поэтому возможно, что столь заметное увеличение активности ряда фенольных соединений, обладающих высокой антиоксидантной активностью, обусловлено защитной реакцией дерева-хозяина на заражение гнилевыми болезнями. Вместе с тем, мы предполагаем, что это одна из дополнительных функций фенольных соединений. Кроме того, обращает внимание ингибирование синтеза фенолгликозидов (арбутина, салицина) в насаждениях, пораженных стволовыми гнилями, детерминирующих параметры индуцированной энтоморезистентности. Полученные результаты предполагают, что развитие гнилевых болезней у березы повислой может сопровождаться снижением уровня энтоморезистентности. Это хорошо соответствует полученным нами ранее результатам о том, что поражение стволовыми гнилями березы сопровождается заметным снижением энтоморезистентности и возрастанием степени дефолиации крон непарным шелкопрядом, по сравнению со здоровыми деревьями (Колтунов, Клобуков, 2009).
Рецензенты:
Усольцев В.А., д.с.-х.н., профессор, профессор Уральского государственного лесотехнического университета, г. Екатеринбург;
Менщиков С.Л., д.с.-х.н., заведующий лабораторией экологии техногенных растительных сообществ ФГБУН Ботанический сад Российской Академии наук, г. Екатеринбург.