



Проблема естественной семенной регенерации дубрав привлекает настойчивое внимание исследователей [1, 2, 3, 4 и др.]. Но роль патогенных организмов в этих процессах остаётся недостаточно изученной. Представляется актуальным устранить в некоторой степени этот пробел и оценить возможность улучшения естественной семенной регенерации дубрав на основе негативного массового отбора на биорезистентность в наиболее уязвимой ювенильной фазе дуба черешчатого.
Негативный массовый отбор широко применяется в лесоводстве. Он заключается в удалении при промежуточных рубках больных, повреждённых, ослабленных деревьев [6].
Цель исследований - оценка эффективности негативного массового отбора на биорезистентность в ювенильной фазе как способа повышения эффективности естественной семенной регенерации свежих дубрав дуба черешчатого.
Объект и методика исследований
Исследования проводились в свежих дубравах дуба черешчатого в предгорьях Северо-Западного Кавказа в период 2001...2006 гг.
Было заложено три блока блоках круговых пробных площадей (КПП), различающихся по уровню композициозного и структурного разнообразия. По другим таксационно-лесоводственным параметрам в пределах каждого блока насаждений допускались лишь минимально возможные различия. Создаваемый ими "информационный шум" компенсировался статистически обоснованным числом оценок исследуемых параметров.
Каждый блок насаждений был предсчтавлен 16-ю КПП. Всего было заложено 48 круговых пробных площадей. Pадиус каждой КПП равнялся 17,84 м, что определяло площадь каждой КПП 1000 м2. Для равномерного охвата исследованием всей круговой пробной площади и статистической обоснованности оценок каждая КПП разбивалась на четыре сектора, ориентированных по странам света (рис. 1).
Учёт естественного возобновления дуба проводился в условиях интервала освещённости 2130 ... 2440 Lx. По нашим наблюдениям, этот уровень освещённости достаточен для нормального развития самосева и подроста дуба черешчатого. Таким образом, недостаток света, как потенциальный фактор снижения жизнеспособности самосева и подроста дуба, был исключён.
Жизнеспособность самосева и подроста дуба оценивалась по 5- балльной шкале: 4-й балл - здоровые особи (без признаков повреждений или ослабленности), 3-й балл - заметно повреждённые или ослабленные особи (степень дефолиации до 20%), 2-й балл - значительно повреждённые или ослабленные особи (степень дефолиации 21 - 50% , 1-й балл - отмирающие особи (степень дефолиации более 50%, 0-й балл - отмершие особи.
Рис. 1. Схема круговой пробной площади
Результаты исследований и их обсуждение
В таблице 1 представлен исходный вариант исследуемых насаждений. В них особи дуба черешчатого, находящиеся в ювенильной фазе развития (самосев и подрост), распределены на пять фенотипов - в зависимости от поражённости насекомыми и грибами. Доминировала мучнистая роса, её конидиальная стадия Oidium dubium Jacz.
Все спорофиты, в том числе и дуб, прежде чем войти в репродуктивную фазу, проходят ювенильное развитие. В ювенильной фазе особи дуба наиболее восприимчивы к инфекционным болезням, особенно к мучнистой росе, поскольку покровные ткани листьев ещё очень нежны, а источник инфекции - поражённые патогеном перезимовавшие листья дуба - находится в непосредственной близости. В крону взрослых деревьев споры мучнистой росы попадают позднее и в меньшем количестве. В ювенильном периоде наиболее чётко проявляется дифференциация молодых особей дуба по резистентности / восприимчивости к инфекционным болезням и другим повреждениям.
Таблица 1. Параметры относительной приспособленности фенотипов дуба черешчатого в ювенильной фазе, различных по степени резистентности (R) / восприимчивости (S) к мучнистой росе (исходный вариант)
|
Параметры относительной приспособленности биотипов |
Биотипы, различные по степени устойчивости/ восприимчивости к мучнистой росе |
||||
|
RR |
R |
RS |
S |
SS |
|
|
Встречаемость особей, % |
2,6 |
23,4 |
32,2 |
26,1 |
15,7 |
|
Относительная приспособленность (f) |
0,08 |
0,73 |
1.0 |
0,81 |
0,49 |
|
Селекционный коэффициент (s) |
0,92 |
0,27 |
0 |
0,19 |
0,51 |
|
Примечание. RR - высокая резистентность, R - повышенная резистентность, RS - популяционный уровень устойчивость/восприимчивость S - повышенная восприимчивость, SS - высокая восприимчивость |
|||||
Как следует из таблицы 1, доминируют (32,2%) в обследованных насаждениях особи самосева и подроста, относящиеся к среднепопуляционному по поражаемости фенотипу RS. На втором месте по встречаемости особи с повышенной к мучнистой росе восприимчивостью (26,1%), на третьем - особи с повышенной резистентностью. Крайние позиции занимают особи с высокой восприимчивостью и высокой резистентностью - соответственно 15,7% и 2,6%. Соответственно этому распределились относительная приспособленность и селекционный коэффициент фенотипов. Имеется в виду, что исследуемые фенотипы являются свободно скрещивающимися, что обычно понимается под менделевской популяцией [5].
В насаждениях каждой круговой пробной площади на двух радиально смежных круговых секторах удалялись SS- и S- особи самосева и подроста дуба, на других смежных парах круговых секторов, принятых за контроль, - нет. Через 3 года учёт особей по степени повреждённости повторился. Результаты повторного учёта представлены в таблицах 2 и 3.
Таблица 2. Параметры относительной приспособленности биотипов дуба черешчатого в ювенильной фазе, различных по степени резистентности (R) / восприимчивости (S) к мучнистой росе (после удаления S- и SS- особей)
|
Параметры относительной приспособ- ленности биотипов |
Биотипы, различные по степени устойчивости/ восприимчивости к мучнистой росе |
||||
|
RR |
R |
RS |
S |
SS |
|
|
Встречаемость, % |
5,2 |
20,4 |
45,0 |
17,2 |
12,2 |
|
Относительная приспособленность (f) |
0,12 |
0,45 |
1.0 |
0,38 |
0,27 |
|
Селекционный коэффициент (s) |
0,88 |
0,55 |
0 |
0,62 |
0,73 |
|
Примечание. RR - высокая резистентность, R - повышенная резистентность, RS - популяционный уровень устойчивость/восприимчивость S - повышенная восприимчивость, SS - высокая восприимчивость |
|||||
Как следует из таблицы 2, в опытном варианте возросла доля биорезистентных фенотипов и уменьшилась доля восприимчивых к болезни фенотипов.
Таблица 3. Параметры относительной приспособленности биотипов дуба черешчатого в ювенильной фазе, различных по степени резистентности (R) / восприимчивости (S) к мучнистой росе (контроль - исходный вариант через 3 года)
|
Параметры относительной приспособленности биотипов |
Биотипы, различные по степени устойчивости/ восприимчивости к мучнистой росе |
||||
|
RR |
R |
RS |
S |
SS |
|
|
Встречаемость, % |
1,1 |
13,4 |
42,0 |
32,1 |
21,7 |
|
Относительная приспособленность (f) |
0,03 |
0,32 |
1.0 |
0,76 |
0,52 |
|
Селекционный коэффициент (s) |
0,97 |
0,68 |
0 |
0,24 |
0,48 |
|
Примечание. RR - высокая резистентность, R - повышенная резистентность, RS - популяционный уровень устойчивость/восприимчивость S - повышенная восприимчивость, SS - высокая восприимчивость |
|||||
Как следует из таблицы 3, в контроле уменьшилась доля биорезистентных фенотипов и увеличилась доля восприимчивых к болезни фенотипов. В таблице 4 (график - рисунок 2) сравниваются селекционные коэффициенты фенотипов в опыте и контроле как критерий эффективности негативного массового отбора на биорезистентность.
Поскольку селекционные коэффициенты количественно характеризуют снижение приспособленности фенотипов в популяции, уменьшение их значений в резистентных фенотипах (RR и R) и увеличение в восприимчивых фенотипах (SS и S) в опытном варианте свидетельствует о положительных изменениях в популяционном составе дуба в ювенильном возрасте в результате массового негативного отбора на биорезистентность.
Таблица 4. Влияние негативного массового отбора на селекционные коэффициенты (s) фенотипов
|
Объект |
Селекционный коэффициент (s) фенотипов |
||||
|
RR |
R |
RS |
S |
SS |
|
|
Опыт |
0,88 |
0,55 |
0 |
0,62 |
0,73 |
|
Контроль |
0,97 |
0,68 |
0 |
0,24 |
0,48 |
|
sо - sк |
- 0,09 |
- 0,13 |
0 |
+ 0,38 |
+ 0,25 |
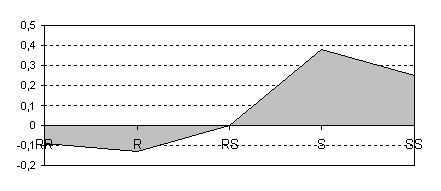
Рис. 2. Влияние негативного массового отбора на селекционный коэффициент резистентных / восприимчивых фенотипов
Следует отметить, что негативный массовый отбор на биорезистентность даёт лишь частичный селекционный выигрыш, поскольку отбор проводится по фенотипу. Решающее значение имеет уровень генетической обусловленности удаляемых минусовых особей дуба. Но в естественных лесах генетическая вариабельность обычно значительна и массовый отбор достаточно эффективен. Кроме того, эффективность массового негативного отбора всегда повышается при его более длительном действии и на большей площади (Rohmeder, Schönbach, 1959). Естественные экологические барьеры, затрудняющие распространение спор патогенных организмов из соседних насаждений, необходимы для получения ожидаемого эффекта от негативного массового отбора на биорезистентность.
Таким образом, негативный массовый отбор на биорезистентность в ювенильной фазе развития дуба черешчатого уменьшает поражаемость самосева и подроста дуба и может служить эффективным способом регуляции патогенеза естественного семенного возобновления во влажных дубравах предгорий Северо-Западного Кавказа.
СПИСОК ЛИТЕРАТУРЫ:
- 1. Демьянов В.П., Солнцев Г.К. // Лесное хозяйство. 1989. № 12. С. 3.
- 2. Зеленко Е.И., Щербань В.А. // Лесное хозяйство. 2000. № 5. С. 3.
- 3. Харченко Н.А., Царалунга В.В. // Вестник Центрально-Чернозёмного регионального отделения наук о лесе РАЕН. Вып. 4, ч. 2. Воронеж, 2002. С. 5.
- 4. Царалунга В.В. // Лесопатологическое и санитарное состояние лесов Воронежской области, пути его оздоровления. Воронеж, ВГУ. 2005. С. 3.
- 5. Высоцкий Н.Л., Высоцкий А.А. // Генетическая оценка исходного материала в лесной селекции. 2000. Воронеж, БИОМИК. С. 15.
- 6. Rohmeder E., Schönbach H. // Hamburg und Berlin, 1959. Verlag Paul Parey. S. 338.